Citation: Jie Wang, Jian Wang, Jiafeng Cao. A heterogeneous parasitic-mutualistic model of mistletoes and birds on a periodically evolving domain[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6678-6698. doi: 10.3934/mbe.2020347

| [1] | J. E. Aukema, C. M. del Rio, Where does a fruit-eating bird deposit mistletoe seeds? seed deposition patterns and an experiment, Ecology, 83 (2002), 3489-3496. |
| [2] |
D. W. Watson, Mistletoe: A keystone resource in forests and woodlands worldwide, Ann. Rev. Ecol. Evol. Syst., 32 (2001), 219-249. doi: 10.1146/annurev.ecolsys.32.081501.114024

|
| [3] |
R. Liu, C. M. del Rio, J. Wu, Spatiotemporal variation of mistletoes: A dynamic modeling approach, Bull. Math. Biol., 73 (2011), 1794-1811. doi: 10.1007/s11538-010-9592-6
|
| [4] | C. C. Wang, R. S. Liu, J. P. Shi, C. M. del Rio, Spatiotemporal mutualistic model of mistletoes and birds, J. Math. Biol., 68 (2014), 1479-1520. |
| [5] | C. C. Wang, R. S. Liu, J. P. Shi, C. M. del Rio, Traveling waves of a mutualistic model of mistletoes and birds, Discrete Contin. Dyn. Syst., 35 (2014), 1743-1765. |
| [6] |
E. J. Crampin, E. A. Gaffney, P. K. Maini, Reaction and diffusion on growing domains: scenarios for robust pattern formation, Bull. Math. Biol., 61 (1999), 1093-1120. doi: 10.1006/bulm.1999.0131
|
| [7] |
E. J. Crampin, E. A. Gaffney, P. K. Maini, Mode-doubling and tripling in reaction-diffusion patterns on growing domains: a piecewise linear model, J. Math. Biol., 44 (2002), 107-128. doi: 10.1007/s002850100112
|
| [8] |
Q. L. Tang, Z. G. Lin, The asymptotic analysis of an insect dispersal model on a growing domain, J. Math. Anal. Appl., 378 (2011), 649-656. doi: 10.1016/j.jmaa.2011.01.057
|
| [9] |
Q. L. Tang, L. Zhang, Z. G. Lin, Asymptotic profile of species migrating on a growing habitat, Acta Appl. Math., 116 (2011), 227-235. doi: 10.1007/s10440-011-9639-1
|
| [10] |
D. H. Jiang, Z. C. Wang, The diffusive logistic equation on periodically evolving domains, J. Math. Anal. Appl., 458 (2018), 93-111. doi: 10.1016/j.jmaa.2017.08.059
|
| [11] | M. Y. Zhang, Z. G. Lin, The diffusive model for Aedes Aegypti mosquito on a periodically evolving domain, Discrete Contin. Dyn. Syst. Ser. B, 24 (2018), 4703. |
| [12] | M. Zhu, Y. Xu, J. D. Cao, The asymptotic profile of a dengue fever model on a periodically evolving domain, Appl. Math. Comput., 362 (2019), 124531. |
| [13] | D. Acheson, Elementary Fluid Dynamics, Oxford University Press, New York, 1990. |
| [14] | M. J. Baines, Moving Finite Element, Monographs on Numerical Analysis, Clarendon Press, Oxford, 1994. |
| [15] | D. Daners, P. K. Medina, Abstract Evolution Equations, Periodic Problems and Applications, Longman Scientific & Technical, Harlow, 1992. |
| [16] | P. Hess, Periodic-Parabolic Boundary Value Problems and Positivity, Pitman Research Notes in Mathematics vol 247, Longman Scientific & Technical, Harlow, 1991. |
| [17] | R. S. Cantrell, C. Cosner, Spatial Ecology via Reaction-Diffusion Equations, John Wiley & Sons Ltd., 2003. |
| [18] |
J. Ge, C. X. Lei, Z. G. Lin, Reproduction numbers and the expanding fronts for a diffusionadvection SIS model in heterogeneous time-periodic environment, Nonlin. Anal. Real World Appl., 33 (2017), 100-120. doi: 10.1016/j.nonrwa.2016.06.005
|
| [19] |
M. Y. Zhang, Z. G. Lin, A reaction-diffusion-advection model for Aedes aegypti mosquitoes in a time-periodic environment, Nonlin. Anal. Real World Appl., 46 (2019), 219-237. doi: 10.1016/j.nonrwa.2018.09.014
|
| [20] |
X. Liang, L. Zhang, X. Q. Zhao, Basic reproduction ratios for periodic abstract functional differential equations (with application to a spatial model for Lyme disease), J. Dynam. Differ. Equat., 31 (2019), 1247-1278. doi: 10.1007/s10884-017-9601-7
|
| [21] |
R. Peng, X. Q. Zhao, A reaction-diffusion SIS epidemic model in a time-periodic environment, Nonlinearity, 25 (2012), 1451-1471. doi: 10.1088/0951-7715/25/5/1451
|
| [22] |
L. Q. Pu, Z. G. Lin, A diffusive SIS epidemic model in a heterogeneous and periodically evolving environment, Math. Biosci. Eng., 16 (2019), 3094-3110. doi: 10.3934/mbe.2019153
|
| [23] |
C. V. Pao, Stability and attractivity of periodic solution parabolic system with time delays, J. Math. Anal. Appl., 304 (2005), 423-450. doi: 10.1016/j.jmaa.2004.09.014
|
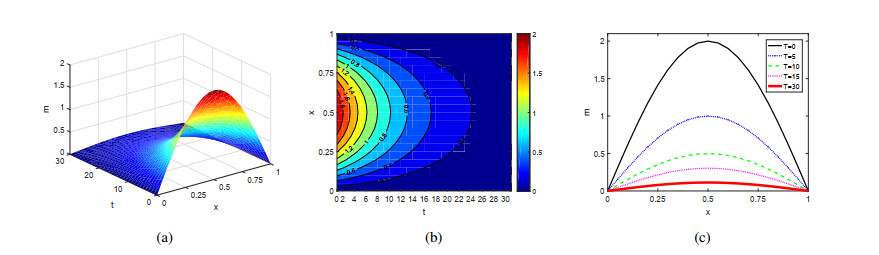
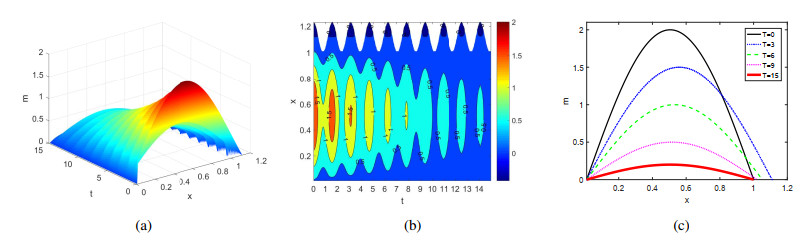
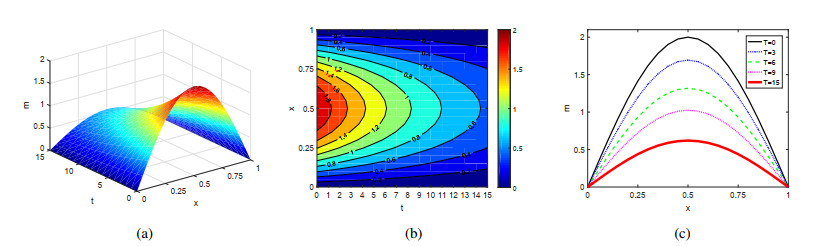
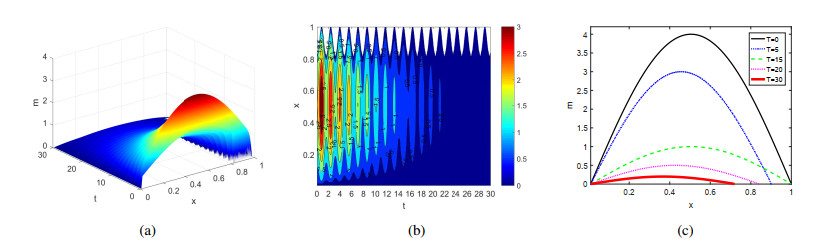
Figures(4)

Jie Wang, Jian Wang, Jiafeng Cao. A heterogeneous parasitic-mutualistic model of mistletoes and birds on a periodically evolving domain[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6678-6698. doi: 10.3934/mbe.2020347







 DownLoad:
DownLoad: