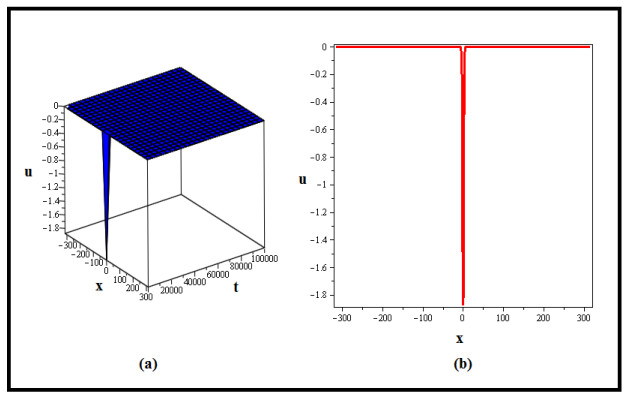
The current work investigates solitary wave solutions for the fractional modified Degasperis-Procesi equation and the fractional gas dynamics equation with Caputo's derivative by using a modified extended direct algebraic method. This method transforms the targeted fractional partial differential equations (FPDEs) into more manageable nonlinear ordinary differential equations, which are then turned into systems of nonlinear algebraic equations with a series-based solution assumption. Using Maple 13, the solitary wave solutions are then obtained by solving the obtained systems. The method produces multiple innovative solitary wave solutions for both equations, which are graphically depicted as 3D and 2D graphs and provide important insights into their behaviors. These insights help us to comprehend wave behavior and the physical processes represented by these equations. Furthermore, the suggested technique exhibits dependability and efficacy in dealing with complicated FPDEs, which bodes well for future studies on the subject.
Citation: Saima Noor, Azzh Saad Alshehry, Asfandyar Khan, Imran Khan. Innovative approach for developing solitary wave solutions for the fractional modified partial differential equations[J]. AIMS Mathematics, 2023, 8(11): 27775-27819. doi: 10.3934/math.20231422
The current work investigates solitary wave solutions for the fractional modified Degasperis-Procesi equation and the fractional gas dynamics equation with Caputo's derivative by using a modified extended direct algebraic method. This method transforms the targeted fractional partial differential equations (FPDEs) into more manageable nonlinear ordinary differential equations, which are then turned into systems of nonlinear algebraic equations with a series-based solution assumption. Using Maple 13, the solitary wave solutions are then obtained by solving the obtained systems. The method produces multiple innovative solitary wave solutions for both equations, which are graphically depicted as 3D and 2D graphs and provide important insights into their behaviors. These insights help us to comprehend wave behavior and the physical processes represented by these equations. Furthermore, the suggested technique exhibits dependability and efficacy in dealing with complicated FPDEs, which bodes well for future studies on the subject.

| [1] |
H. Y. Jin, Z. A. Wang, L. Wu, Global dynamics of a three-species spatial food chain model, J. Differ. Equ., 333 (2022), 144–183. https://doi.org/10.1016/j.jde.2022.06.007 doi: 10.1016/j.jde.2022.06.007

|
| [2] |
P. Liu, J. Shi, Z. A. Wang, Pattern formation of the attraction-repulsion Keller-Segel system, Discrete Cont. Dyn. S., 18 (2013), 2597–2625. https://doi.org/10.3934/dcdsb.2013.18.2597 doi: 10.3934/dcdsb.2013.18.2597
|
| [3] |
H. Khan, R. Shah, P. Kumam, M. Arif, Analytical solutions of fractional-order heat and wave equations by the natural transform decomposition method, Entropy, 21 (2022), 597. https://doi.org/10.3390/e21060597 doi: 10.3390/e21060597
|
| [4] | H. Y. Jin, Z. A. Wang, Global stabilization of the full attraction-repulsion Keller-Segel system, arXiv Preprint, 2019. |
| [5] |
L. Liu, S. Zhang, L. Zhang, G. Pan, J. Yu, Multi-UUV maneuvering counter-game for dynamic target scenario based on fractional-order recurrent neural network, IEEE T. Cybernetics, 53 (2022), 4015–4028. https://doi.org/10.1109/TCYB.2022.3225106 doi: 10.1109/TCYB.2022.3225106
|
| [6] |
A. M. Zidan, A. Khan, M. K. Alaoui, W. Weera, Evaluation of time-fractional Fishers equations with the help of analytical methods, AIMS Math., 7 (2022), 18746–18766. https://doi.org/10.3934/math.20221031 doi: 10.3934/math.20221031
|
| [7] |
R. Luo, Z. Peng, J. Hu, On model identification based optimal control and it's applications to multi-agent learning and control, Mathematics, 11 (2023), 906. https://doi.org/10.3390/math11040906 doi: 10.3390/math11040906
|
| [8] |
X. Lyu, X. Wang, C. Qi, R. Sun, Characteristics of cavity dynamics, forces, and trajectories on vertical water entries with two spheres side-by-side, Phys. Fluids, 35 (2023). https://doi.org/10.1063/5.0166794 doi: 10.1063/5.0166794
|
| [9] |
E. M. Elsayed, R. Shah, K. Nonlaopon, The analysis of the fractional-order Navier-Stokes equations by a novel approach, J. Funct. Space., 2022 (2022), 1–18. https://doi.org/10.1155/2022/8979447 doi: 10.1155/2022/8979447
|
| [10] |
M. Ekici, A. Sonmezoglu, E. M. Zayed, A new fractional sub-equation method for solving the space-time fractional differential equations in mathematical physics, Comput. Methods Diffe., 2 (2014), 153–170. https://doi.org/10.9734/BJMCS/2013/2908 doi: 10.9734/BJMCS/2013/2908
|
| [11] |
H. Khan, R. Shah, J. F. G. Aguilar, D. Baleanu, P. Kumam, Travelling waves solution for fractional-order biological population model, Math. Model. Nat. Pheno., 16 (2021), 32. https://doi.org/10.1051/mmnp/2021016 doi: 10.1051/mmnp/2021016
|
| [12] |
A. Ara, N. A. Khan, O. A. Razzaq, T. Hameed, M. A. Z. Raja, Wavelets optimization method for evaluation of fractional partial differential equations: An application to financial modelling, Adv. Differ. Equ., 2018 (2018), 1–13. https://doi.org/10.1186/s13662-017-1461-2 doi: 10.1186/s13662-017-1461-2
|
| [13] |
H. Sun, Y. Zhang, D. Baleanu, W. Chen, Y. Chen, A new collection of real world applications of fractional calculus in science and engineering, Commun. Nonlinear Sci., 64 (2018), 213–231. https://doi.org/10.1016/j.cnsns.2018.04.019 doi: 10.1016/j.cnsns.2018.04.019
|
| [14] |
P. Sunthrayuth, N. H. Aljahdaly, A. Ali, I. Mahariq, A. M. Tchalla, $\psi$-Haar wavelet operational matrix method for fractional relaxation-oscillation equations containing $\psi$-Caputo fractional derivative, J. Funct. Space., 2021 (2021), 1–14. https://doi.org/10.1155/2021/7117064 doi: 10.1155/2021/7117064
|
| [15] |
M. K. Alaoui, K. Nonlaopon, A. M. Zidan, A. Khan, Analytical investigation of fractional-order cahn-hilliard and gardner equations using two novel techniques, Mathematics, 10 (2022), 1643. https://doi.org/10.3390/math10101643 doi: 10.3390/math10101643
|
| [16] |
N. J. Ford, J. Xiao, Y. Yan, A finite element method for time fractional partial differential equations, Fract. Calc. Appl. Anal., 14 (2011), 454–474. https://doi.org/10.2478/s13540-011-0028-2 doi: 10.2478/s13540-011-0028-2
|
| [17] |
D. Fulger, E. Scalas, G. Germano, Monte Carlo simulation of uncoupled continuous-time random walks yielding a stochastic solution of the space-time fractional diffusion equation, Phys. Rev. E, 77 (2008), 021122. https://doi.org/10.1103/PhysRevE.77.021122 doi: 10.1103/PhysRevE.77.021122
|
| [18] |
A. R. Alharbi, M. B. Almatrafi, Exact solitary wave and numerical solutions for geophysical KdV equation, J. King Saud Univ. Sci., 34 (2022), 102087. https://doi.org/10.1016/j.jksus.2022.102087 doi: 10.1016/j.jksus.2022.102087
|
| [19] |
M. B. Almatrafi, A. Alharbi, K. Lotfy, A. A. El-Bary, Exact and numerical solutions for the GBBM equation using an adaptive moving mesh method, Alex. Eng. J., 60 (2021), 4441–4450. https://doi.org/10.1016/j.aej.2021.03.023 doi: 10.1016/j.aej.2021.03.023
|
| [20] |
A. R. Alharbi, M. B. Almatrafi, M. A. Abdelrahman, Analytical and numerical investigation for Kadomtsev-Petviashvili equation arising in plasma physics, Phys. Scripta, 95 (2020), 045215. https://doi.org/10.1088/1402-4896/ab6ce4 doi: 10.1088/1402-4896/ab6ce4
|
| [21] |
M. A. Abdelrahman, M. B. Almatrafi, A. Alharbi, Fundamental solutions for the coupled KdV system and its stability, Symmetry, 12 (2020), 429. https://doi.org/10.3390/sym12030429 doi: 10.3390/sym12030429
|
| [22] |
A. R. Alharbi, M. B. Almatrafi, Analytical and numerical solutions for the variant Boussinseq equations, J. Taibah Univ. Sci., 14 (2020), 454–462. https://doi.org/10.1080/16583655.2020.1746575 doi: 10.1080/16583655.2020.1746575
|
| [23] |
A. M. Zidan, A. Khan, M. K. Alaoui, W. Weera, Evaluation of time-fractional Fishers equations with the help of analytical methods, AIMS Math., 7 (2022), 18746–18766. https://doi.org/10.3934/math.20221031 doi: 10.3934/math.20221031
|
| [24] |
K. Xu, Y. Guo, Y. Liu, X. Deng, Q. Chen, Z. Ma, 60-GHz compact dual-mode on-chip bandpass filter using GaAs technology, IEEE Electr. Device L., 42 (2021), 1120–1123. https://doi.org/10.1109/LED.2021.3091277 doi: 10.1109/LED.2021.3091277
|
| [25] |
Z. Li, K. Wang, W. Li, S. Yan, F. Chen, S. Peng, Analysis of surface pressure pulsation characteristics of centrifugal pump magnetic liquid sealing film, Front. Energy Res., 10 (2022), 937299. https://doi.org/10.3389/fenrg.2022.937299 doi: 10.3389/fenrg.2022.937299
|
| [26] |
H. Chen, W. Chen, X. Liu, X. Liu, Establishing the first hidden-charm pentaquark with strangeness, Eur. Phys. J. C, 81 (2021), 409. https://doi.org/10.1140/epjc/s10052-021-09196-4 doi: 10.1140/epjc/s10052-021-09196-4
|
| [27] |
M. Ayata, O. Ozkan, A new application of conformable Laplace decomposition method for fractional Newell-Whitehead-Segel equation, AIMS Math., 5 (2020), 7402–7412. https://doi.org/10.3934/math.2020474 doi: 10.3934/math.2020474
|
| [28] |
H. Khan, D. Baleanu, P. Kumam, J. F. Al-Zaidy, Families of travelling waves solutions for fractional-order extended shallow water wave equations, using an innovative analytical method, IEEE Access, 7 (2019), 107523–107532. https://doi.org/10.1109/ACCESS.2019.2933188 doi: 10.1109/ACCESS.2019.2933188
|
| [29] |
B. Q. Li, Y. L. Ma, Rich soliton structures for the Kraenkel-Manna-Merle (KMM) system in ferromagnetic materials, J. Supercond. Nov. Magn., 31 (2018), 1773–1778. https://doi.org/10.1007/s10948-017-4406-9 doi: 10.1007/s10948-017-4406-9
|
| [30] |
B. Q. Li, Y. L. Ma, The non-traveling wave solutions and novel fractal soliton for the (2+1)-dimensional Broer–Kaup equations with variable coefficients, Commun. Nonlinear Sci., 16 (2011), 144–149. https://doi.org/10.1016/j.cnsns.2010.02.011 doi: 10.1016/j.cnsns.2010.02.011
|
| [31] |
S. Mukhtar, S. Noor, The numerical investigation of a fractional-order multi-dimensional Model of Navier-Stokes equation via novel techniques, Symmetry, 14 (2022), 1102. https://doi.org/10.3390/sym14061102 doi: 10.3390/sym14061102
|
| [32] |
S. Duran, Exact solutions for time-fractional Ramani and Jimbo-Miwa equations by direct algebraic method, Adv. Sci. Eng. Medicine, 12 (2020), 982–988. https://doi.org/10.1166/asem.2020.2663 doi: 10.1166/asem.2020.2663
|
| [33] |
R. Y. Molliq, M. S. M. Noorani, I. Hashim, R. R. Ahmad, Approximate solutions of fractional Zakharov-Kuznetsov equations by VIM, J. Comput. Appl. Math., 233 (2009), 103–108. https://doi.org/10.1016/j.cam.2009.03.010 doi: 10.1016/j.cam.2009.03.010
|
| [34] |
B. Zheng, Exp-function method for solving fractional partial differential equations, The Scientific World J., 2013 (2013). https://doi.org/10.1186/1687-1847-2013-199 doi: 10.1186/1687-1847-2013-199
|
| [35] |
Y. L. Ma, B. Q. Li, Y. Y. Fu, A series of the solutions for the Heisenberg ferromagnetic spin chain equation, Math. Method. Appl. Sci., 41 (2018), 3316–3322. https://doi.org/10.1002/mma.4818 doi: 10.1002/mma.4818
|
| [36] |
Y. L. Ma, B. Q. Li, C. Wang, A series of abundant exact travelling wave solutions for a modified generalized Vakhnenko equation using auxiliary equation method, Appl. Math. Comput., 211 (2009), 102–107. https://doi.org/10.1016/j.amc.2009.01.036 doi: 10.1016/j.amc.2009.01.036
|
| [37] |
B. Q. Li, Y. L. Ma, Periodic solutions and solitons to two complex short pulse (CSP) equations in optical fiber, Optik, 144 (2017), 149–155. https://doi.org/10.1016/j.ijleo.2017.06.114 doi: 10.1016/j.ijleo.2017.06.114
|
| [38] |
M. Zhang, Y. L. Ma, B. Q. Li, Novel loop-like solitons for the generalized Vakhnenko equation, Chinese Phys. B, 22 (2013), 030511. https://doi.org/10.1088/1674-1056/22/3/030511 doi: 10.1088/1674-1056/22/3/030511
|
| [39] |
Y. L. Ma, B. Q. Li, Novel optical soliton structures for a defocusing Lakshmanan-Porsezian-Daniel optical system, Optik, 284 (2023), 170931. https://doi.org/10.1016/j.ijleo.2023.170931 doi: 10.1016/j.ijleo.2023.170931
|
| [40] |
B. Q. Li, Y. L. Ma, Hybrid soliton and breather waves, solution molecules and breather molecules of a (3+1)-dimensional Geng equation in shallow water waves, Phys. Lett. A, 463 (2023), 128672. https://doi.org/10.1016/j.physleta.2023.128672 doi: 10.1016/j.physleta.2023.128672
|
| [41] |
H. Yasmin, N. H. Aljahdaly, A. M. Saeed, R. Shah, Probing families of optical soliton solutions in fractional perturbed Radhakrishnan-Kundu-Lakshmanan model with improved versions of extended direct algebraic method, Fractal Fract., 7 (2023), 512. https://doi.org/10.3390/fractalfract7070512 doi: 10.3390/fractalfract7070512
|
| [42] |
Q. Al-Mdallal, K. A. Abro, I. Khan, Analytical solutions of fractional Walter's B fluid with applications, Complexity, 2018 (2018), 1–10. https://doi.org/10.1155/2018/8131329 doi: 10.1155/2018/8131329
|
| [43] |
H. Jiang, F. Liu, I. Turner, K. Burrage, Analytical solutions for the multi-term time-fractional diffusion-wave/diffusion equations in a finite domain, Comput. Math. Appl., 64 (2012), 3377–3388. https://doi.org/10.1016/j.camwa.2012.02.042 doi: 10.1016/j.camwa.2012.02.042
|
| [44] |
B. Zheng, Exp-function method for solving fractional partial differential equations, The Scientific World J., 2013 (2013). https://doi.org/10.1186/1687-1847-2013-199 doi: 10.1186/1687-1847-2013-199
|
| [45] |
H. Khan, S. Barak, P. Kumam, M. Arif, Analytical solutions of fractional Klein-Gordon and gas dynamics equations, via the (G'/G)-expansion method, Symmetry, 11 (2019), 566. https://doi.org/10.3390/sym11040566 doi: 10.3390/sym11040566
|
| [46] |
Y. Tian, J. Liu, Direct algebraic method for solving fractional Fokas equation, Thermal Sci., 25 (2021), 2235–2244. https://doi.org/10.2298/TSCI200306111T doi: 10.2298/TSCI200306111T
|
| [47] |
H. Gnerhan, F. S. Khodadad, H. Rezazadeh, M. M. Khater, Exact optical solutions of the (2+1) dimensions Kundu-Mukherjee-Naskar model via the new extended direct algebraic method, Mod. Phys. Lett. B, 34 (2020), 2050225. https://doi.org/10.1142/S0217984920502255 doi: 10.1142/S0217984920502255
|
| [48] |
M. Younis, M. Iftikhar, Computational examples of a class of fractional order nonlinear evolution equations using modified extended direct algebraic method, J. Comput. Methods Sci., 15 (2015), 359–365. https://doi.org/10.3233/JCM-150548 doi: 10.3233/JCM-150548
|
| [49] |
H. Yasmin, N. H. Aljahdaly, A. M. Saeed, R. Shah, Investigating symmetric soliton solutions for the fractional coupled Konno-Onno system using improved versions of a novel analytical technique, Mathematics, 11 (2023), 2686. https://doi.org/10.3390/math11122686 doi: 10.3390/math11122686
|
| [50] |
H. Yasmin, N. H. Aljahdaly, A. M. Saeed, R. Shah, Investigating families of soliton solutions for the complex structured coupled fractional Biswas-Arshed model in birefringent fibers using a novel analytical technique, Fractal Fract., 7 (2023), 491. https://doi.org/10.3390/fractalfract7070491 doi: 10.3390/fractalfract7070491
|
| [51] |
T. A. A. Ali, Z. Xiao, H. Jiang, B. Li, A class of digital integrators based on trigonometric quadrature rules, IEEE T. Ind. Electron., 2023. https://doi.org/10.1109/TIE.2023.3290247 doi: 10.1109/TIE.2023.3290247
|
| [52] |
Y. Fang, H. Min, X. Wu, W. Wang, X. Zhao, G. Mao, On-ramp merging strategies of connected and automated vehicles considering communication delay, IEEE Intel. Transp. Sy., 23 (2022), 15298–15312. https://doi.org/10.1109/TITS.2022.3140219 doi: 10.1109/TITS.2022.3140219
|
| [53] |
W. Kuang, H. Wang, X. Li, J. Zhang, Q. Zhou, Y. Zhao, Application of the thermodynamic extremal principle to diffusion-controlled phase transformations in Fe-C-X alloys: Modeling and applications, Acta Mater., 159, (2018), 16–30. https://doi.org/10.1016/j.actamat.2018.08.008 doi: 10.1016/j.actamat.2018.08.008
|
| [54] |
A. M. Wazwaz, New solitary wave solutions to the modified forms of Degasperis-Procesi and Camassa-Holm equations, Appl. Mathe. Comput., 186 (2007), 130–141. https://doi.org/10.1016/j.amc.2006.07.092 doi: 10.1016/j.amc.2006.07.092
|
| [55] |
D. Li, S. S. Ge, T. H. Lee, Fixed-Time-Synchronized consensus control of multiagent systems, IEEE T. Control Netw., 8 (2021), 89–98. https://doi.org/10.1109/TCNS.2020.3034523 doi: 10.1109/TCNS.2020.3034523
|
| [56] |
Q. Meng, Q. Ma, Y. Shi, Adaptive Fixed-time stabilization for a class of uncertain nonlinear systems, IEEE T. Automat. Contr., 2023 (2023). https://doi.org/10.1109/TAC.2023.3244151 doi: 10.1109/TAC.2023.3244151
|
| [57] |
H. Yuan, B. Yang, System dynamics approach for evaluating the interconnection performance of Cross-Border transport infrastructure, J. Manag. Eng., 38 (2022). https://doi.org/10.1061/(ASCE)ME.1943-5479.0001015 doi: 10.1061/(ASCE)ME.1943-5479.0001015
|
| [58] |
V. P. Dubey, R. Kumar, J. Singh, D. Kumar, An efficient computational technique for time-fractional modified Degasperis-Procesi equation arising in propagation of nonlinear dispersive waves, J. Ocean Eng. Sci., 6 (2021), 30–39. https://doi.org/10.1016/j.joes.2020.04.006 doi: 10.1016/j.joes.2020.04.006
|
| [59] |
K. Zhang, A. S. Alshehry, N. H. Aljahdaly, R. Shah, N. A. Shah, M. R. Ali, Efficient computational approaches for fractional-order Degasperis-Procesi and Camassa-Holm equations, Results Phys., 50 (2023), 106549. https://doi.org/10.1016/j.rinp.2023.106549 doi: 10.1016/j.rinp.2023.106549
|
| [60] |
S. Das, R. Kumar, Approximate analytical solutions of fractional gas dynamic equations, Appl. Math. Comput., 217 (2011), 9905–9915. https://doi.org/10.1016/j.amc.2011.03.144 doi: 10.1016/j.amc.2011.03.144
|
| [61] | M. A. Dokuyucu, Caputo and Atangana-Baleanu-Caputo fractional derivative applied to garden equation, Turk. J. Sci., 5 (2020), 1–7. |
| [62] |
S. M. M. Alizamini, H. Rezazadeh, K. Srinivasa, A. Bekir, New closed form solutions of the new coupled Konno-Oono equation using the new extended direct algebraic method, Pramana, 94 (2020), 1–12. https://doi.org/10.1007/s12043-020-1921-1 doi: 10.1007/s12043-020-1921-1
|

Figures(2)

Saima Noor, Azzh Saad Alshehry, Asfandyar Khan, Imran Khan. Innovative approach for developing solitary wave solutions for the fractional modified partial differential equations[J]. AIMS Mathematics, 2023, 8(11): 27775-27819. doi: 10.3934/math.20231422







 DownLoad:
DownLoad: