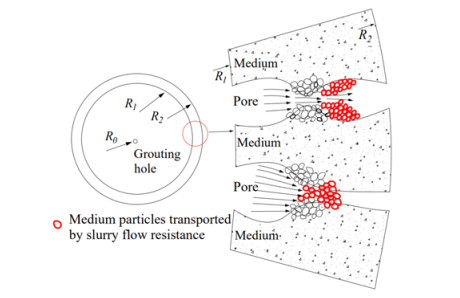
Grouting is widely used to solve problems such as water inrush and collapse in tunnels and subways, with permeation grouting being the most common method. This study explores the influence of fluid flow resistance on the permeability of porous media and derives an analytic expression for the variation of permeability in porous media based on the diffusion law of cement-based grout columns-hemispherical diffusion. Based on this, a Newtonian fluid permeation grouting column-hemispherical diffusion model considering the variation of permeability in porous media is established, combining the rheological equation and the equilibrium equation of the Newtonian fluid. The theoretical results considering the influence of fluid flow resistance on the permeability of porous media are consistent with the results of existing laboratory tests and numerical simulations, which are closer to the experimental results, thus verifying the effectiveness of the proposed theoretical mechanism. Parameter analysis shows that with the increase of grouting pressure and grouting time, the diffusion radius of the Newtonian fluid infiltration grouting, considering the variation of permeability in porous media, increases rapidly and then increases slowly, and with the increase of porous media porosity, its radius decreases gradually. The research results can provide theoretical support for selecting grouting parameters and quality control in porous media infiltration grouting engineering practice.
Citation: Qi Peng, Shaobo Yang, Guangcan Shan, Nan Qiao. Analysis of the diffusion range of the sleeve valve pipe permeation grouting column-hemispheric under the variation of the permeability of porous media[J]. Electronic Research Archive, 2023, 31(11): 6928-6946. doi: 10.3934/era.2023351
Grouting is widely used to solve problems such as water inrush and collapse in tunnels and subways, with permeation grouting being the most common method. This study explores the influence of fluid flow resistance on the permeability of porous media and derives an analytic expression for the variation of permeability in porous media based on the diffusion law of cement-based grout columns-hemispherical diffusion. Based on this, a Newtonian fluid permeation grouting column-hemispherical diffusion model considering the variation of permeability in porous media is established, combining the rheological equation and the equilibrium equation of the Newtonian fluid. The theoretical results considering the influence of fluid flow resistance on the permeability of porous media are consistent with the results of existing laboratory tests and numerical simulations, which are closer to the experimental results, thus verifying the effectiveness of the proposed theoretical mechanism. Parameter analysis shows that with the increase of grouting pressure and grouting time, the diffusion radius of the Newtonian fluid infiltration grouting, considering the variation of permeability in porous media, increases rapidly and then increases slowly, and with the increase of porous media porosity, its radius decreases gradually. The research results can provide theoretical support for selecting grouting parameters and quality control in porous media infiltration grouting engineering practice.

| [1] | G. Zhu, Q. Zhang, X. Feng, R. T. Liu, L. Z. Zhang, S. Q. Liu, Study on the filtration mechanism in permeation grouting using the particle deposition probability model, Adv. Eng. Sci., 52 (2020), 125–135. |
| [2] |
Z. Sun, D. Zhang, Y. Hou, N. Huangfu, M. Li, F. Guo, Support countermeasures for large deformation in a deep tunnel in layered shale with high geostresses, Rock Mech. Rock Eng., 56 (2023), 4463–4484. https://doi.org/10.1007/s00603-023-03297-0 doi: 10.1007/s00603-023-03297-0

|
| [3] |
Z. Sun, D. Zhang, Q. Fang, N. Huangfu, Z. Chu, Convergence-confinement analysis for tunnels with combined bolt–cable system considering the effects of intermediate principal stress, Acta Geotech., 18 (2023), 3323–3348. https://doi.org/10.1007/s11440-022-01758-4 doi: 10.1007/s11440-022-01758-4
|
| [4] |
Z. Sun, D. Zhang, Q. Fang, D. Liu, G. Dui, Displacement process analysis of deep tunnels with grouted rock bolts considering bolt installation time and bolt length, Comput. Geotech., 140 (2021), 104437. https://doi.org/10.1016/j.compgeo.2021.104437 doi: 10.1016/j.compgeo.2021.104437
|
| [5] | J. Wang, Geotechnical Grouting Theory and Engineering Examples, Beijing: Science Press, (2001), 101–105. |
| [6] |
D. Gouvenot, State of the art in European grouting, Proc. Inst. Civ. Eng. Ground Improv., 2 (1998), 51–67. https://doi.org/10.1680/gi.1998.020201 doi: 10.1680/gi.1998.020201
|
| [7] | K. Wang, Mechanism of Grouting Reinforcement for Completely Weathered Granite with Rich Water and Its Application, PhD thesis, Shandong University, 2017. |
| [8] | S. Li, W. Zhao, Y. Huang, Y. Lei, L. Yu, Study on the characteristics of grout permeation based on cylindrical diffusion, J. Coal Sci. Eng. (China), 19 (2013), 57–62. |
| [9] |
C. Han, J. Wei, W. Zhang, F. Yang, H. Yin, D. Xie, et al., Quantitative permeation grouting in sand layer with consideration of grout properties and medium characteristics, Constr. Build. Mater., 327 (2022), 126947. https://doi.org/10.1016/j.conbuildmat.2022.126947 doi: 10.1016/j.conbuildmat.2022.126947
|
| [10] |
F. Celik, O. Akcuru, Rheological and workability effects of bottom ash usage as a mineral additive on the cement based permeation grouting method, Constr. Build. Mater., 263 (2020), 120186. https://doi.org/10.1016/j.conbuildmat.2020.120186 doi: 10.1016/j.conbuildmat.2020.120186
|
| [11] |
L. G. Baltazar, F. M. A. Henriques, F. Jorne, Optimisation of flow behaviour and stability of superplasticized fresh hydraulic lime grouts through design of experiments, Constr. Build. Mater., 35 (2012), 838–845. https://doi.org/10.1016/j.conbuildmat.2012.04.084 doi: 10.1016/j.conbuildmat.2012.04.084
|
| [12] | C. Zhang, J. Yang, G. Zhang, X. Ye, Y. Zhang, Z. Zhang, et al., Experiment and application research on stability performance of filling grouting slurry, Chin. J. Rock Mech. Eng., 37 (2018), 3604–3612. |
| [13] |
J. Zhou, J. Lu, Y. Zhang, Z. Wang, Calculation formula of permeation grouting considering slurry viscosity variation, J. Mater. Sci. Eng., 37 (2019), 758–762. https://doi.org/10.14136/j.cnki.issn1673-2812.2019.05.013 doi: 10.14136/j.cnki.issn1673-2812.2019.05.013
|
| [14] | S. Li, Research on Permeation Grouting Experiment and Application in Sandy Strata, PhD thesis, Northeastern University, 2010. |
| [15] | Q. Lu, Z. Q. Yang, Z. Q. Yang, R. X. Yu, Y. Y. Zhu, Y. Yang, Penetration grouting mechanism of Bingham fluid considering diffusion paths, Rock Soil Mech., 43 (2022), 385–394. |
| [16] | Y. Wang, Study on Infiltration Grouting Mechanism of Newtonian Fluid Based on Tortuosity of Porous Media, PhD thesis, Kunming University of Science and Technology, 2020. |
| [17] | Z. Yang, K. Hou, T. T. Guo, Q. Ma, Study of column-hemispherical penetration grouting mechanism based on Bingham fluid of time-dependent behavior of viscosity, Rock Soil Mech., 32 (2011), 2697–2703. |
| [18] | F. Ye, C. Gou, Z. Chen, Y. Liu, J. Zhang, Back-filled grouts diffusion model of shield tunnel considering its viscosity degeneration, China J. Highway Transp., 26 (2013), 127–134. |
| [19] | L. Z. Zhang, Q. S. Zhang, R. T. Liu, S. C. Li, H. B. Wang, W. Li, Penetration grouting mechanism of quick setting slurry considering spatiotemporal variation of viscosity, Rock Soil Mech., 38 (2017), 443–452. |
| [20] | K. Fang, T. Xia, L. Bao, J. Ye, S. Chen, Cement grout dispersion considering filtration in gravel, Chin. J. Rock Mech. Eng., 32 (2013), 1443–1448. |
| [21] | S. Li, Z. Zheng, R. Liu, X. Feng, Z. Sun, L. Zhang, Analysis of diffusion of grout in porous media considering infiltration effects, Chin. J. Rock Mech. Eng., 34 (2015), 2401–2409. |
| [22] | S. Li, X. Feng, R. Liu, L. Zhang, W. Han, Z. Zheng, Diffusion of grouting cement in sandy soil considering filtration effect, Rock Soil Mech., 38 (2017), 925–933. |
| [23] | Y. Fu, X. Wang, S. Zhang, Y. Yang, Modelling of permeation grouting considering grout self-gravity effect: theoretical and experimental study, Adv. Mater. Sci. Eng., 2019 (2019). https://doi.org/10.1155/2019/7968240 |
| [24] | W. Yang, Study on Permeation Grouting Mechanism of Porous Media Based on Grout Self-Gravity, PhD thesis, Kunming University of Science and Technology, 2020. |
| [25] | M. Yuan, Study on Some Flow Characteristics of Fluid in Porous Media, PhD thesis, Huazhong University of Science and Technology, 2008. |
| [26] | F. Ren, Study on Flow Resistance of Porous Media Based on Variable Diameter Tube Ball Mode, PhD thesis, Inner Mongolia University of Science and Technology, 2022. |
| [27] |
B. Sun, X. Li, Z. Wang, B. Ma, H. He, Laboratory investigation of the effect of the pore pressure on argillaceous siltstone permeability, Eng. Geol., 317 (2023), 107067. https://doi.org/10.1016/j.enggeo.2023.107067 doi: 10.1016/j.enggeo.2023.107067
|
| [28] |
H. Qu, Y. Peng, J. Huang, Z. Pan, F. Zhou, Modelling of the impact of stress concentration on permeability in porous medium based on machine learning method, Geoenergy Sci. Eng., 224 (2023), 211655. https://doi.org/10.1016/j.geoen.2023.211655 doi: 10.1016/j.geoen.2023.211655
|
| [29] |
I. Palmer, J. Mansoori, How permeability depends on stress and pore pressure in coalbeds: a new model, SPE Reservoir Eval. Eng., 1 (1998), 539–544. https://doi.org/10.2118/52607-PA doi: 10.2118/52607-PA
|
| [30] | L. Zhang, M. Tian, S. Lu, M. Li, J. Li, Analysis of permeability variation and stress sensitivity of liquid nitrogen fracturing coal with different water contents, Rock Soil Mech., 43 (2022), 107–116. |
| [31] | F. Sun, J. Yang, M. Wang, B. Sun, J. Zhang, Y. Zhao, et al., Stress sensitivity characteristics and control mechanism of different coal rank reservoirs, Coal Geol. Explor., 50 (2022), 51–58. |
| [32] | J Bear, Dynamics of Fluids in Porous Media, American Elsevier Publishing Company, INC. (1972), 129–131. |
| [33] | A. S. Iberall, Permeability of glass wool and other highly porous media, J. Res. Natl. Bur. Stand., 45 (1950), 398–406. |
| [34] |
D. R. E. Harleman, P. F. Mehlhorn, R. R. Rumer, Dispersion-permeability correlation in porous, J. Hydraul. Div., 89 (1963), 67–85. https://doi.org/10.1061/JYCEAJ.0000863 doi: 10.1061/JYCEAJ.0000863
|
| [35] | Z. Liu, S. Dong, H. Wang, Law of grout diffusion of horizontal hole grouting in incline crack, J. China Coal Soc., 47 (2022), 135–151. |
| [36] | X. Kong, Higher Seepage Mechanics, China University of Science and Technology Press, 1999. |
| [37] | Z. Yang, K. Hou, W. Liang, Y. Cheng, B. Yang, Study of diffusion parameters of Newtonian fluid based on column-hemispherical penetration grouting, Rock Soil Mech., 35 (2014), 17–24. |
| [38] | W. Ruan, Research on diffusion of grouting and basic properties of grouts, Chin. J. Geotech. Eng., 27 (2005), 69–73. |
| [39] | Y. Wang, A numerical simulation of the permeation grouting process in metro construction, Natl. Def. Transp. Eng. Technol., 14 (2016), 42–44. |
| [40] | Z. Yang, X. Niu, K. Hou, Y. Guo, W. Liang, Z. Zhou, Column penetration grouting mechanism researches based on Power-law fluid, J. Harbin Inst. Technol., 48 (2016), 178–183. |
| [41] | S. Maghous, Z. Saada, L. Dormieux, J. Canou, J. C. Dupla, A model for in situ grouting with account for particle filtration, Comput. Geotech., 34 (2007), 164–174. |









Figures(10) / Tables(5)
Qi Peng, Shaobo Yang, Guangcan Shan, Nan Qiao. Analysis of the diffusion range of the sleeve valve pipe permeation grouting column-hemispheric under the variation of the permeability of porous media[J]. Electronic Research Archive, 2023, 31(11): 6928-6946. doi: 10.3934/era.2023351







 DownLoad:
DownLoad: