Citation: Ross Sadler. Towards a more complete understanding of the occurrence and toxicities of the cylindrospermopsins[J]. AIMS Environmental Science, 2015, 2(3): 827-851. doi: 10.3934/environsci.2015.3.827
| [1] | Zhun Han, Hal L. Smith . Bacteriophage-resistant and bacteriophage-sensitive bacteria in a chemostat. Mathematical Biosciences and Engineering, 2012, 9(4): 737-765. doi: 10.3934/mbe.2012.9.737 |
| [2] | Baojun Song, Wen Du, Jie Lou . Different types of backward bifurcations due to density-dependent treatments. Mathematical Biosciences and Engineering, 2013, 10(5&6): 1651-1668. doi: 10.3934/mbe.2013.10.1651 |
| [3] | Timothy C. Reluga, Jan Medlock . Resistance mechanisms matter in SIR models. Mathematical Biosciences and Engineering, 2007, 4(3): 553-563. doi: 10.3934/mbe.2007.4.553 |
| [4] | Linda J. S. Allen, P. van den Driessche . Stochastic epidemic models with a backward bifurcation. Mathematical Biosciences and Engineering, 2006, 3(3): 445-458. doi: 10.3934/mbe.2006.3.445 |
| [5] | Miller Cerón Gómez, Eduardo Ibarguen Mondragon, Eddy Lopez Molano, Arsenio Hidalgo-Troya, Maria A. Mármol-Martínez, Deisy Lorena Guerrero-Ceballos, Mario A. Pantoja, Camilo Paz-García, Jenny Gómez-Arrieta, Mariela Burbano-Rosero . Mathematical model of interaction Escherichia coli and Coliphages. Mathematical Biosciences and Engineering, 2023, 20(6): 9712-9727. doi: 10.3934/mbe.2023426 |
| [6] | Kento Okuwa, Hisashi Inaba, Toshikazu Kuniya . An age-structured epidemic model with boosting and waning of immune status. Mathematical Biosciences and Engineering, 2021, 18(5): 5707-5736. doi: 10.3934/mbe.2021289 |
| [7] | Lingli Zhou, Fengqing Fu, Yao Wang, Ling Yang . Interlocked feedback loops balance the adaptive immune response. Mathematical Biosciences and Engineering, 2022, 19(4): 4084-4100. doi: 10.3934/mbe.2022188 |
| [8] | Olga Vasilyeva, Tamer Oraby, Frithjof Lutscher . Aggregation and environmental transmission in chronic wasting disease. Mathematical Biosciences and Engineering, 2015, 12(1): 209-231. doi: 10.3934/mbe.2015.12.209 |
| [9] | Maoxing Liu, Yuming Chen . An SIRS model with differential susceptibility and infectivity on uncorrelated networks. Mathematical Biosciences and Engineering, 2015, 12(3): 415-429. doi: 10.3934/mbe.2015.12.415 |
| [10] | G. V. R. K. Vithanage, Hsiu-Chuan Wei, Sophia R-J Jang . Bistability in a model of tumor-immune system interactions with an oncolytic viral therapy. Mathematical Biosciences and Engineering, 2022, 19(2): 1559-1587. doi: 10.3934/mbe.2022072 |
Bacteriophage or virulent phage is a virus which can grow and replicate by infecting bacteria. Once residing in bacteria, phage grow quickly, which result in the infection of bacteria and drive the bacteria to die [17]. Thus, we can view them as bacteria predators and use them to cure the diseases induced by infecting bacteria [9,35]. Phage therapy has become a promising method because of the emergence of antibiotic resistant bacteria [12,9]. Indeed, as a treatment, phages have several advantages over antibiotics. Phages replicate and grow exponentially, while antibiotics are not [47]. Generally speaking, a kind of phages infect only particular classes of bacteria, and this limitation of their host is very beneficial to cure the diseases. Moreover, phages are non-toxic, and cannot infect human cells. Hence, there are fewer side effects as compared to antibiotics [12,35].
It is important to understand the interaction dynamics between bacteria and phage to design an optimal scheme of phage therapy. There have been a number of papers that study the mathematical models of bacteriophages (see, for example, [1,2,3,4,8,9,10,11,21,23,28,42] and the references cited therein). Campbell [11] proposed a deterministic mathematical model for bacteria and phage, which is a system of differential equations containing two state variables, susceptible bacteria
It was observed that phage can exert pressure on bacteria to make them produce resistance through loss or modification of the receptor molecule to which a phage binds with an inferior competition ability for nutrient uptake [25]. More recently, biological evidences are found that there exists an adaptive immune system across bacteria, which is the Clustered regularly interspaced short palindromic repeats (CRISPRs) along with Cas proteins [6,14,13,18,26,27,31,41]. In this system, phage infection is memorized via a short invader sequence, called a proto-spacer, and added into the CRISPR locus of the host genome. And, the CRISPR/Cas system admits heritable immunity [14,13,18,26,31]. The replication of infecting phage in bacteria is aborted if their DNA matches the crRNAs (CRISPR RNAs) which contains these proto-spacer. On the other hand, if there is no perfect pairing between the proto-spacer and the foreign DNA (as in the case of a phage mutant), the CRISPR/Cas system is counteracted and replication of the phage DNA can occur [19,34,33,39]. Therefore, the CRISPR/Cas system participates in a constant evolutionary battle between phage and bacteria [7,13,27,47].
Mathematical models are powerful in understanding the population dynamics of bacteria and phages. Han and Smith [23] formulated a mathematical model that includes a phage-resistant bacteria, where the resistant bacteria is an inferior competitor for nutrient. Their analytical results provide a set of sufficient conditions for the phage-resistant bacteria to persist. Recently, mathematical models have been proposed to study the contributions of adaptive immune response from CRISPR/Cas in bacteria and phage coevolution [27,29]. In these papers, numerical simulations are used to find how the immune response affects the coexistence of sensitive strain and resistance strain of bacteria. In the present paper, we extend the model in [23] by incorporating the CRISPR/Cas immunity on phage dynamics. Following [23], we focus on five state variables:
Let
|
$
˙R=D(R0−R)−f(R)(S+μM),˙S=−DS+f(R)S−kSP,˙M=−DM+μf(R)M+εkSP−k′MP,˙I=−DI+(1−ε)kSP+k′MP−δI,˙P=−DP−kSP−kMP+bδI,
$
|
(1) |
where a dot denotes the differentiation with respect to time
The paper is organized as follows. In the next section we present the mathematical analysis of the model that include the stability and bifurcation of equilibria. Numerical simulations are provided in Section
In this section, we present the mathematical analysis for the stability and bifurcations of equilibria of (1). We start with the positivity and boundedness of solutions.
Proposition 1. All solutions of model (1) with nonnegative initial values are nonnegative. In particular, a solution
Proof. We examine only the last conclusion of the proposition. First, we claim that
| $S(t)=S(0)e^{\int_0^t[-D-kP(\theta)+f(R(\theta))]d\theta}>0.$ |
In a similar way we can show the positivity of
Proposition 2. All nonnegative solutions of model (1) are ultimately bounded.
Proof. Set
| $L(t)=R(t)+S(t)+M(t)+I(t)+\frac{1}{b}P(t).$ |
Calculating the derivative of
|
$
˙L(t)=DR0−DR(t)−DS(t)−DM(t)−DI(t)−1bDP(t) −1bkS(t)P(t)−1bkM(t)P(t)≤DR0−DL(t).
$
|
It follows that the nonnegative solutions of
| $ \label{upperlimit} \limsup\limits_{t\rightarrow\infty}L(t)\leq R^0. $ | (2) |
Therefore, the nonnegative solutions of model (1) are ultimately bounded.
| $ m>D, \;\;f(R^0)>D. $ | (3) |
Then
| $ \mu m>D, \;\;\mu f(R^0)>D. $ | (4) |
It is easy to see that (3) and (4) imply
| $\lambda_1<\mu\lambda_2<\lambda_2.$ |
Thus, the competitive exclusion in the absence of phage infection holds [24], and the boundary equilibrium
The basic reproduction number
|
$F=\left(
0(1−ε)k(R0−λ1)00
\right),
$
|
and
|
$V=\left(
D+δ0−bδD+k(R0−λ1)
\right),
$
|
and is defined as the spectral radius of
| $R_0=\sqrt\frac{b\delta(1-\varepsilon)k(R^0-\lambda_1)}{(D+\delta)[D+k(R^0-\lambda_1)]}.$ |
Analogously, the basic reproduction number
| $R^{M}_0=\sqrt\frac{b\delta\kappa(R^0-\lambda_2)}{(D+\delta)[D+k(R^0-\lambda_2)]}.$ |
Theorem 2.1. The infection-free equilibrium
The proof of Theorem 2.1 is postponed to Appendix.
Theorem 2.2. The infection-free equilibrium
Proof. Define a Lyapunov function by
| $V(t)=R(t)-R_1-\int_{R_1}^R\frac{f(R_1)}{f(\xi)}d\xi+S(t)-S_1-S_1\ln\frac{S}{S_1}+M(t)+I(t)+\frac{1}{b}P(t), $ |
where
|
$
˙V(t)=(1−f(R1)f(R))˙R(t)+(1−S1S)˙S(t)+˙M(t)+˙I(t)+1b˙P(t)=D(R0−R)−D(R0−R)f(R1)f(R)+(S+μM)f(R1)+(D+kP−f(R))S1−DS−DM−DI−DbP−kSP+kMPb=D(R0−R1)(2−f(R1)f(R)−f(R)f(R1))−D(R−R1)(1−f(R1)f(R))−D(1−μ)M−DI+(kS1−Db)P−kSP+kMPb.
$
|
Since
| $ D_0=\{(R, S, M, I, P)\mid \dot{V}=0\}. $ |
It is easy to examine that the largest invariant set in
| $ \{(R, S, M, I, P)\mid R=\lambda_1, S=S_1, M=0, I=0, P=0\}. $ |
It follow from the LaSalle's invariance principle [22] that
In this subsection, we consider the infection equilibria of system (1) which satisfy
|
$
D(R0−R)−f(R)S−μf(R)M=0,−DS+f(R)S−kSP=0,−DM+μf(R)M+εkSP−k′MP=0,−DI+(1−ε)kSP+k′MP−δI=0,−DP−kSP−kMP+bδI=0.
$
|
(5) |
If
|
$
D(R0−R3)−μf(R3)M3=0,−DM3+μf(R3)M3−k′M3P3=0,−DI3+k′M3P3−δI3=0,−DP3−kM3P3+bδI3=0.
$
|
It follows that
| $P_3=\frac{\mu f(R_3)-D}{k'}, \quad M_3=\frac{D(R^0-R_3)}{\mu f(R_3)}, \quad I_3=\frac{k'M_3P_3}{D+\delta}, $ |
and
|
$
−DP3−kM3P3+bδI3=P3(kA2M3−D)=0,
$
|
where
| $ g(R_3):=R_3^2+(a+mA-R^0)R_3-R^0 a=0, $ | (6) |
where
| $ A=\frac{\mu}{k}\frac{D+\delta}{b\delta\kappa-(D+\delta)}. $ |
By direct calculations, we obtain
| $ g(R^0)=mAR^0>0, \\ g(\lambda_2)=(\lambda_2-R^0)(\lambda_2+a)+mA\lambda_2. $ |
Since
| $A<\frac{(R^0-\lambda_2)(\lambda_2+a)}{m\lambda_2} =\frac{\mu(R^0-\lambda_2)}{D}, $ |
which is equivalent to
Theorem 2.3. The infection equilibrium
To study the local stability of infection equilibria
|
$
a1=kM3+3D+δ+μM3f′(R3),a2=kM3(μM3f′(R3)+2D−μf(R3))+D(2D+δ)+(2D+δ+μf(R3))μM3f′(R3),a3=DkM3(μM3f′(R3)+D−μf(R3))+D(D+δ)(μf(R3)−D)+μf(R3)μM3f′(R3)(2D+δ),a4=kA2(D+δ)(μM3f′(R3)+D)(μf(R3)−D).
$
|
(7) |
Moreover, for
| $ \label{L3} \lambda_3=f^{-1}(D(1-\kappa)/(\mu-\kappa)). $ | (8) |
Theorem 2.4. The infection-resistant equilibrium
| $ \kappa<\mu, \;\;R_3>\lambda_3, \;\;a_1a_2>a_3, \;\;(a_1a_2-a_3)a_3-a_4a_1^2>0, $ | (9) |
and is unstable when
| $ \kappa<\mu, \;\;R_3<\lambda_3, \;\;a_1a_2>a_3, \;\;(a_1a_2-a_3)a_3-a_4a_1^2>0. $ | (10) |
The proof of Theorem 2.4 is given in Appendix.
Let us now consider the existence of coexistence equilibrium of (1). Denote such an equilibrium by
|
$
P4=f(R4)−Dk,M4=εD(R0−R4)(f(R4)−D)[D−μf(R4)+με(f(R4)−D)+k′P4]f(R4),S4=D(R0−R4)[k′P4+D−μf(R4)][D−μf(R4)+με(f(R4)−D)+k′P4]f(R4),I4=[(1−ε)kS4+k′M4]P4D+δ.
$
|
(11) |
Since
| $ \label{Positive} f(R_4)>D, \;\; k'P_4+D-\mu f(R_4)>0. $ | (12) |
Note that
|
$
F(R4):=k′P4+D−μf(R4)=D(1−κ)+(κ−μ)f(R4),
$
|
where
Note that
| $ -D+\left(\frac{b\delta\kappa}{D+\delta}-1\right)kM_4+\left(b\delta \frac{1-\varepsilon}{D+\delta}-1\right)kS_4=0. $ | (13) |
Set
| $ A_1=b\delta(1-\varepsilon)/(D+\delta)-1, \quad A_2=b\delta\kappa/(D+\delta)-1. $ | (14) |
Using (11) and
| $ G(R_4):=k(R^0-R_4)\left(e_1+e_2\frac{D}{f(R_4)}\right)-(e_3f(R_4)+e_4)=0, $ |
where
|
$
e1=κA1−μA1+εA2,e2=A1−κA1−εA2,e3=με−μ+κ,e4=(1−με−κ)D.
$
|
Let
Evidently,
| $ G(\lambda_2)=-D+kA_2(R^0-\lambda_2)=(D+k(R^0-\lambda_2))(R^{M}-1). $ |
Thus,
| $ G(\lambda_1)=-D+kA_1(R^0-\lambda_1)=(D+k(R^0-\lambda_1))(R_0^2-1). $ |
Hence,
For the case where
|
$
G(λ3)=D(R0−λ3)(e1+e2Df(λ3))−(e3f(λ3)+e4)=D(R0−λ3)εA2(1−μ)1−κ−μεD(1−μ)μ−κ.
$
|
Since
| $ \lambda_3\geq R^0-\frac{\mu(1-\kappa)}{A_2(\mu-\kappa)}, $ | (15) |
and
| $ \lambda_3<R^0-\frac{\mu(1-\kappa)}{A_2(\mu-\kappa)}. $ | (16) |
Solve
|
$
b∗=D+δδ((μ−κ)(R0−λ3)(1−κ)μ+1).
$
|
Notice that
|
$
P4=f(λ3)−Dk=D(1−κ)k(μ−κ),M4=D(R0−λ3)(f(λ3)−D)μf(λ3),I4=k′M4P4D+δ.
$
|
Set
|
$
R∗:=R0RM0=√(D+k(R0−λ2))(1−ε)(R0−λ1)κ(R0−λ2)(D+k(R0−λ1)).
$
|
Solving
| $ \varepsilon=\varepsilon^{*}:=1-\frac{\kappa(R^0-\lambda_2)(D+k(R^0-\lambda_1))}{(D+k(R^0-\lambda_2))(R^0-\lambda_1)}. $ |
Thus,
Let us consider three cases:
The following Theorem states the existence of infection equilibria of (1) according to the above discussions.
Theorem 2.5. Let
For
Proof. Note that
This theorem indicates that the system (1) exhibits a backward bifurcation as
Notice that the equilibrium
Theorem 2.6. Let
For
For
The proof of this Theorem is omitted because it is similar to it for Theorem 2.5.
Theorem 2.6 presents the conditions for a forward bifurcation of the infection-free equilibrium and a transcritical bifurcation of the coexist equilibrium. Note that for
We now explore the persistence and extinction of phages in the case where
Theorem 2.7. Let
(
| $ \lim\limits_{t\to\infty}(R(t), S(t), M(t), I(t), P(t))=(\lambda_1, R^0-\lambda_1, 0, 0, 0) $ |
if
(
| $ \liminf\limits_{t\to \infty} S(t)>\eta, \;\; \liminf\limits_{t\to \infty} M(t)>\eta, \;\; \liminf\limits_{t\to \infty} I(t)>\eta, \;\;\liminf\limits_{t\to \infty} P(t)>\eta. $ |
Proof.
| $ f^{\infty}=\limsup\limits_{t\to \infty}f(t), \;\;f_{\infty}=\liminf\limits_{t\to \infty}f(t). $ |
First, we claim that a positive solution of (1) admits
| $ \dot{S}\leq -D S+f(R)S\leq -D S+f(R^{0}+\eta_0-S)S, \;\;\text{for all large } t, $ |
where
| $ \dot{I}\leq -DI+(1-\varepsilon)k(R^{0}-\lambda_1+\eta_1)P-\delta I, \\ \dot{P}\leq -D P-k(R^{0}-\lambda_1-\eta_1)P+b\delta I, $ | (17) |
where
| $ \dot{I}=-DI+(1-\varepsilon)k(R^{0}-\lambda_1+\eta_1)P-\delta I, \\ \dot{P}=-D P-k(R^{0}-\lambda_1-\eta_1)P+b\delta I. $ | (18) |
The Jacobian matrix of (18) is
|
$
J_1:=\left(
−(D+δ)(1−ε)k(R0−λ1+η1)bδ−(D+k(R0−λ1−η1)
\right).
$
|
Since
|
$
˙R=D(R0−R)−f(R)(S+μM),˙S=−DS+f(R)S,˙M=−DM+μf(R)M.
$
|
(19) |
Since
|
$
X={(R,S,M,I,P):R≥0,S≥0,M≥0,I≥0,P≥0},X0={(R,S,M,I,P)∈X:I>0,P>0},∂X0=X∖X0.
$
|
We wish to show that (1) is uniformly persistent with respect to
By Proposition 1, we see that both
| $ J_{\partial}=\{(R, S, M, I, P)\in X: I=0, P=0\}. $ |
It is clear that there are three equilibria
Note that (3) and (4) imply that a positive solution of (1) cannot approach
| $ \dot{I}\geq -DI+(1-\varepsilon)k(R^{0}-\lambda_1-\eta_2)P-\delta I, \\ \dot{P}\geq -D P-k(R^{0}-\lambda_1+\eta_2)P+b\delta I, $ | (20) |
where
|
$
J_2:=\left(
−(D+δ)(1−ε)k(R0−λ1−η2)bδ−(D+k(R0−λ1+η2)
\right).
$
|
Since
| $ \dot{I}=-DI+(1-\varepsilon)k(R^{0}-\lambda_1-\eta_2)P-\delta I, \\ \dot{P}=-D P-k(R^{0}-\lambda_1+\eta_2)P+b\delta I $ | (21) |
tend to infinity as
By adopting the same techniques as above, we can show that the population
| $ \dot{M}\geq -D M+\varepsilon kSP. $ |
This, together with the uniform persistence of population
In this section, we implement numerical simulations to illustrate the theoretical results and explore more interesting solution patterns of model (1). Take the same parameter values as those in [23] where
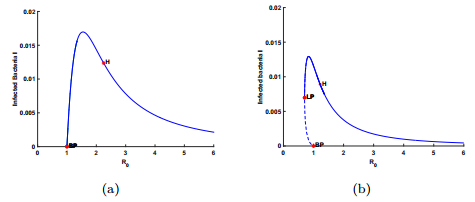 Figure 1. Bifurcation graphs for
Figure 1. Bifurcation graphs for To demonstrate the second case where
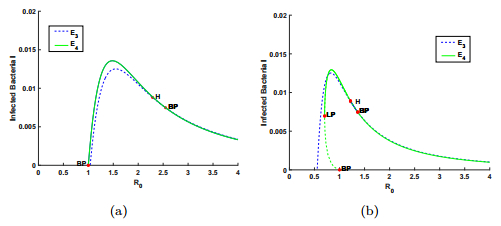 Figure 2. Bifurcation graphs for case
Figure 2. Bifurcation graphs for case To show the case
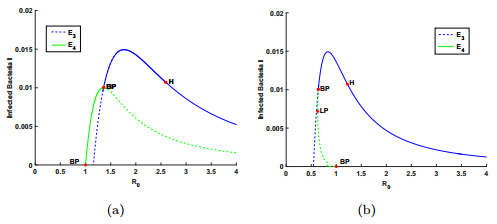 Figure 3. Bifurcation graphs for case
Figure 3. Bifurcation graphs for case With the help of
As discussed above, a bistable coexistence between the infection-free equilibrium and an infection equilibrium may occur when
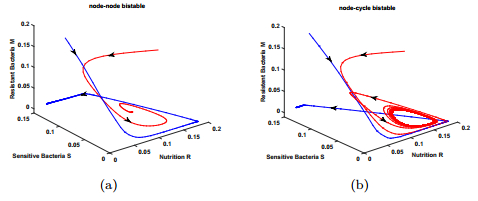 Figure 4. Graphs of bistable behaviors in case
Figure 4. Graphs of bistable behaviors in case In this paper, we have developed a bacteriophage mathematical model based on CRISPR/Cas immune system. By combining theoretical analysis and numerical simulations, we have found that the model exhibits some new dynamical behaviors than the model without the immune responses in [23]. More specifically, the introduction of the CRISPR/Cas immune system induces a backward bifurcation from the infection-free equilibrium or a transcritical bifurcation from the coexist equilibrium, which means that although the basic infection reproduction number is below unity, the phage could coexist with bacteria. The coexistence of a stable infection-free equilibrium with a stable infection equilibrium(or stable coexist equilibrium), the bistable phenomenon of a stable infection-free equilibrium and a stable periodic solution are found, which are shown in panel (a) of Fig. 4 and panel (b) of Fig. 4. They provide reasonable explanations for the complexity of phage therapy [9,29] or bacteria-phages coevolution [13], and the coexistence of bacteria with phage in the biological experiments [20,29]. In contrast, there is no the backward bifurcation or bistable phenomena in the models of previous studies [23,42] where the immune response is ignored.
For case
The mathematical analysis for the stability and bifurcation of equilibria of (1) in this paper present some insights into the underlying phage infection mechanisms by considering the CRISPR/Cas system in bacteria. It will be interesting to consider the analytical conditions for the Hopf bifurcation and the homoclinic bifurcation of the model and reveal how the immune response affect these bifurcations. It will be also interesting to consider the effect of latent period of infection like it in [23] or the nonlinear death rates like those in [29]. We leave these as future researches.
We are very grateful to the anonymous referees for careful reading and valuable comments which have led to important improvements of our original manuscript.
Proof Theorem 2.1. Let
|
$J(E_1)=\left(
−D−S1f′(λ1)−D−μD00S1f′(λ1)000−kS100−D+μD0εkS1000−D−δ(1−ε)kS1000bδ−D−kS1
\right),
$
|
where
| $ (\omega+D)(\omega+S_1f^{'}(\lambda_1))[\omega^2+(2D+\delta+kS_1) \omega+f_0](\omega+D(1-\mu))=0, $ |
where
| $ f_0=(D+kS_1)(D+\delta)(1-R_0^2). $ |
Since
Proof Theorem 2.4. Evaluating the Jacobian of (1) at
|
$J(E_3)=\left(
−D−μM3f′(R3)−f(R3)−μf(R3)000−D+f(R3)−kP3000μM3f′(R3)εkP300−k′M30(1−ε)kP3k′P3−D−δk′M30−kP3−kP3bδ−D−kM3
\right).
$
|
Using
| $(\omega-f(R_3)+kP_3+D)(\omega^4+a_1\omega^3+a_2\omega^2+a_3\omega+a_4)=0, $ |
where
| $ F_1(\omega)=\omega-f(R_3)+kP_3+D, \;\;F_2(\omega)=\omega^4+a_1\omega^3+a_2\omega^2+a_3\omega+a_4. $ |
Note that
| $ f(R_3)-kP_3-D=(1-\frac{\mu}{\kappa})f(R_3)+(\frac{1}{\kappa}-1)D:=F_0(\omega).\nonumber $ |
It is easy to see
| $ f(R_3)>\frac{D(1-\kappa)}{\mu-\kappa}, \nonumber $ |
which is equivalent to
| [1] | Hawkins PR, Runnegar MTC, Jackson ARB, et al. (1985) Severe hepatotoxicity caused by the tropical cyanobacterium (bluegreen alga) Cylindrospermopsis racoborskii (Woloszynska) Seenaya and Subba Raju isolaged from a domestic water supply reservoir. Appl Environ Microb 50: 1292-1295. |
| [2] |
Ohtani I, Moore RE, Runnegar MTC (1992) Cylindrospermopsin: a potent hepatotoxin from the blue-green alga Cylindrospermopsis raciborskii. J Am Chem Soc 114: 7941-7942. doi: 10.1021/ja00046a067

|
| [3] | Seawright AA, Nolan CC, Shaw GR, et al. (1999) The oral toxicity for mice of the tropical cyanobacterium Cylindrospermopsis raciborskii (Woloszynska). Environ Toxicol 14: 135-142. |
| [4] | Humpage A (2008) Toxin types, toxicokinetics and toxicodynamics. Adv Exp Med Biol619: 383-416. |
| [5] |
Kinnear S (2010) Cylindrospermopsin: A Decade of Progress on Bioaccumulation Research. Mar Drugs 8: 542-564. doi: 10.3390/md8030542
|
| [6] | Moreira C, Azevedo J, Antunes A, et al. (2013) Cylindrospermopsin: occurrence, methods of detection and toxicology. J Appl Microb 114: 605-620. |
| [7] | de la Cruz AA, Hiskia A, Kaloudis T, et al. (2013) A review on cylindrospermopsin: the global occurrence, detection, toxicity and degradation of a potent cyanotoxin. Environmental Sci Proc Impacts 15: 1979-2003. |
| [8] | Stewart I, Seawright AA, Schluter PJ, et al. (2006) Primary irritant and delayed-contact hypersensitivity reactions to the freshwater cyanobacterium Cylindrospermopsis raciborskii and its associated toxin cylindrospermopsin. BMC Dermatol 6: PMC1544345. |
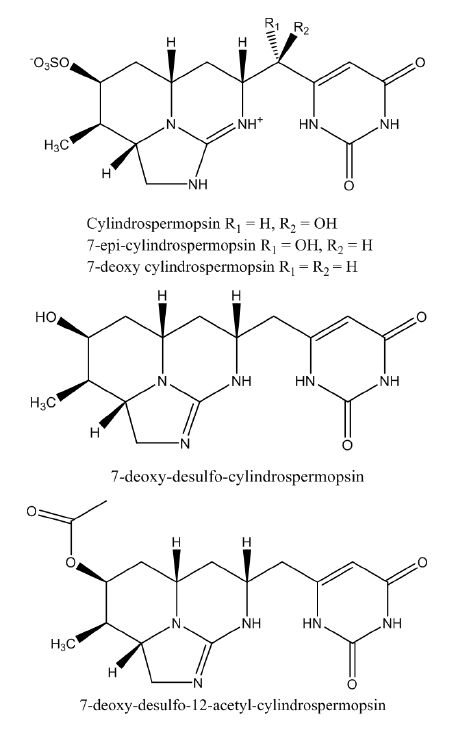
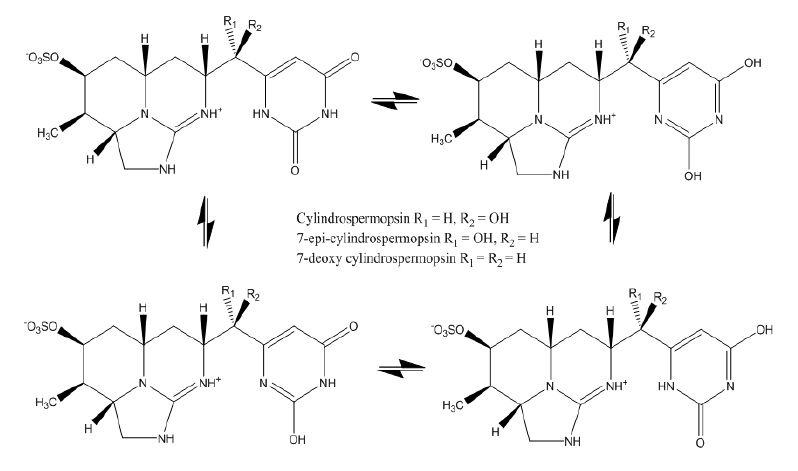
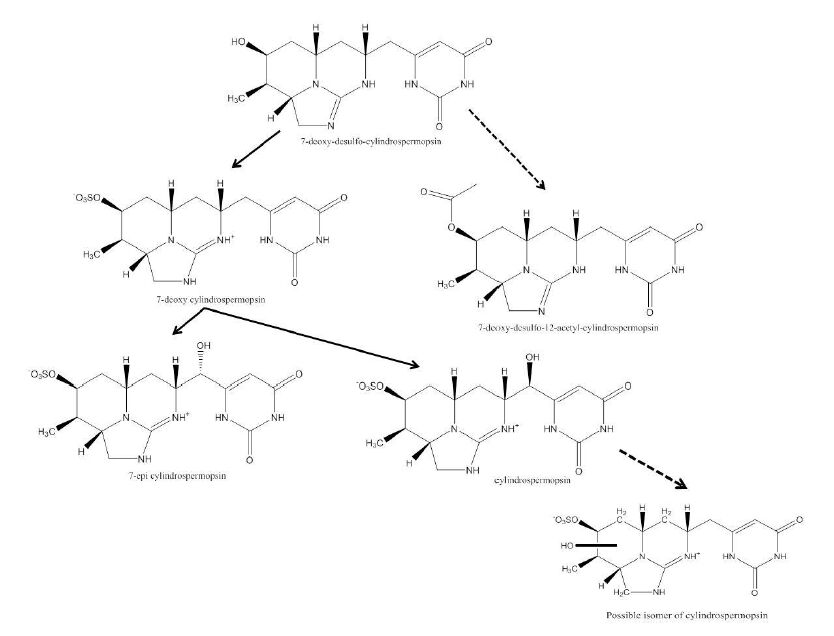
| [9] | Wimmer KM, Strangman WK, Wright JLC (2014) 7-Deoxy-desulfo-cylindrospermopsin and 7-deoxy-desulfo-12-acetylcylindrospermopsin: Two new cylindrospermopsin analogs isolated from a Thai strain of Cylindrospermopsis raciborskii. Harmful Algae 37: 203-206. |
| [10] | Orr PT, Rasmussen JP, Burford MA, et al. (2011) Evaluation of quantitative real-time PCR to characterise spatial and temporal variations in cyanobacteria, Cylindrospermopsis raciborskii (Woloszynska) Seenaya et Subba Raju and cylindrospermopsin concentrations in three subtropical Australian reservoirs. Corrigendum. Harmful Algae 10: 234. |
| [11] |
Li R, Carmichael WW, Brittain S, et al. (2001) First Report of the Cyanotoxins Cylindrospermopsin and Deoxycylindrospermopsin from Raphidiopsis curvata (Cyanobacteria). J Phycol 37: 1121-1126. doi: 10.1046/j.1529-8817.2001.01075.x
|
| [12] | Neumann C, Bain P, Shaw G (2007) Studies of the comparative in vitro toxicology of the cyanobacterial metabolite deoxycylindrospermopsin. J Toxicol Env Health, Part A 70: 1679-1686. |
| [13] |
Everson S, Fabbro L, Kinnear S, et al. (2009) Distribution of the cyanobacterial toxins cylindrospermopsin and deoxycylindrospermopsin in a stratified lake in north-eastern New South Wales, Australia. Mar Fresh Res 60: 25-33. doi: 10.1071/MF08115
|
| [14] |
McGregor G, Sendall BC, Hunt LT, et al. (2011) Report of the cyanotoxinscylindrospermopsin and deoxy-cylindrospermopsin from Raphidiopsis mediterranea Skuja (Cyanobacteria/Nostocales). Harmful Algae 10: 402-410. doi: 10.1016/j.hal.2011.02.002
|
| [15] |
Rzymski P, Poniedziałek B, Kokocin ski M, et al. (2014) Interspecific allelopathy in cyanobacteria: Cylindrospermopsin and Cylindrospermopsis raciborskii effect on the growth and metabolism of Microcystis aeruginosa. Harmful Algae 35: 1-8. doi: 10.1016/j.hal.2014.03.002
|
| [16] |
Heintzelman GR, Fang WK, Keen SP, et al. (2002) Stereoselective total synthesis and reassignment of stereochemistry of the freshwater cyanobacterial hepatotoxins cylindrospermopsin and 7-epi-cylindrospermopsin. J Am Chem Soc 124: 3939-3945. doi: 10.1021/ja020032h
|
| [17] |
Moustaka-Gouni M, Kormas KA, Vardaka E, et al. (2009) Raphidiopsis mediterranea Skuja represents non-heterocytous life-cycle stages of Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju in Lake Kastoria (Greece), its type locality: Evidence by morphological and phylogenetic analysis. Harmful Algae 8: 864-872. doi: 10.1016/j.hal.2009.04.003
|
| [18] |
Banker R, Teltsch B, Sukenik A, et al. (2000) 7-epi-cylindrospermopsin, a toxic minor metabolite of the cyanobacterium Aphanizomenon ovalisporumfrom Lake Kinneret, Israel. J Nat Prod 63: 387-389. doi: 10.1021/np990498m
|
| [19] | Mazmouz R, Chapuis-Hugon F, Pichon V, et al. (2010) Biosynthesis of Cylindrospermopsin and 7-epi-cylindrospermopsin in Oscillatoria sp. Strain PCC 6506: Identification of the cyr Gene Cluster and Toxin Analysis. Appl Environ Microb 76: 4943-4949. |
| [20] | Mazmouz R, Chapuis-Hugon F, Pichon V, et al. (2011) The Last Step of the Biosynthesis of the CyanotoxinsCylindrospermopsin and 7-epi-Cylindrospermopsin is Catalysed by CyrI, a 2-Oxoglutarate- Dependent Iron Oxygenase. Chem Bio Chem 12: 858-862. |
| [21] | Norris RLG, Eaglesham GK, Pierens G, et al. (1999) Deoxycylindropermopsin, an analog of cylindropermopsin from Cylindrospermopsis raciborskii. Environ Toxicol 14: 163-165. |
| [22] | Jiang Y, Xiao P, Yu G, et al. (2012) Molecular Basis and Phylogenetic Implications of Deoxycylindrospermopsin Biosynthesis in the Cyanobacterium Raphidiopsiscurvata, Appl Environ Microb 78: 2256-2263. |
| [23] | Seifert M, McGregor G, Eaglesham G, et al. (2007) First evidence for the production of cylindrospermopsin and deoxycylindrospermopsin by the freshwater benthic cyanobacterium, Lyngbyawollei (Farlow ex Gomont) Speziale and Dyck, JHarmful Algae 6: 73-80. |
| [24] | Evans DM, Murphy PJ (2011) Chapter 1—TheCylindrospermopsin Alkaloids,Elsevier.The Alkaloids: Chemistry and Biology 70: 1-77. |
| [25] |
Looper RE, Runnegar MTC, Williams RM (2006) Syntheses of the cylindrospermopsin alkaloids. Tetrahedron 62: 4549-4562. doi: 10.1016/j.tet.2006.02.044
|
| [26] |
Ríos V, Prieto AI, Cameán AM, et al. (2014) Detection of cylindrospermopsin toxin markers in cyanobacterial algal blooms using analytical pyrolysis (Py-GC/MS) and thermally-assisted hydrolysis and methylation (TCh-GC/MS). Chemosphere 108: 175-182. doi: 10.1016/j.chemosphere.2014.01.033
|
| [27] | Orr PT, Rasmussen JP, Burford MA, et al. (2011) Evaluation of quantitative real-time PCR to characterize spatial and temporal variations in cyanobacteria, Cylindrospermopsis raciborskii (Woloszynska) Seenaya et Subba Raju and cylindrospermopsin concentrations in three subtropical Australian reservoirs. Harmful Algae 9: 243-254. |
| [28] | Shihana F, Jayasekera JMKB, Dissananyake DM, et al. (2012) The short term effect of cyanobacterial toxin extracts on mice kidney. In: Symposium Proceedings, International Symposium on Water Quality and Human Health: Challenges Ahead, PGIS, Peradeniya, Sri Lanka. |
| [29] |
Davis TW, Orr PT, Boyer GL, et al. (2014) Investigating the production and release of cylindrospermopsin and deoxy-cylindrospermopsin by Cylindrospermopsis raciborskii over a natural growth cycle. Harmful Algae 31: 18-25. doi: 10.1016/j.hal.2013.09.007
|
| [30] | Dissananyake DM, Jayasekera JMKB, Ratnayake P, et al. (2012) Effect of Concentrated Water from Reservoirs of high prevalence Area for Chronic Kidney Disease (CKDu) of unknown Origin in Sri Lanka on Mice, In: Symposium Proceedings, International Symposium on Water Quality and Human Health: Challenges Ahead, PGIS, Peradeniya, Sri Lanka. |
| [31] | Saker ML, Griffiths DJ (2000) The Effect of Temperature on Growth and Cylindrospermopsiin Content of seven isolates of Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) from Water bodies in Northern Australia. Phycologia 39: 349-354. |
| [32] |
Bácsi I, Vasas G, Surányi, G, et al. (2006) Alteration of cylindrospermopsin production in sulfate- or phosphate-starved cyanobacterium Aphanizomenon ovalisporum. FEMS Microbiol Lett 259: 303-310. doi: 10.1111/j.1574-6968.2006.00282.x
|
| [33] | Preussel K, Wessel G, Fastner J, et al. (2009) Response of cylindrospermopsin production and release in Aphanizomenon flos-aquae (Cyanobacteria) to varying light and temperature conditions. Harmful Algae 6: 645-650. |
| [34] |
Dyble J, Tester PA, Litaker RW (2006) Effects of light intensity on cylindrospermopsin production in the cyanobacterial HAB species Cylindrospermopsis raciborskii. Afr J Mar Sci 28: 309-312. doi: 10.2989/18142320609504168
|
| [35] | Chiswell RK, Shaw GR, Eaglesham G, et al. (1999) Stability of cylindrospermopsin, the toxin from the cyanobacterium, Cylindrospermopsis raciborskii: effect of pH, temperature, and sunlight on decomposition. Environ Toxicol 14: 155-161. |
| [36] | Smith M, Shaw GR, Eaglesham, GK, et al. (2008) Elucidating the Factors Influencing the Biodegradation of Cylindrospermopsin in Drinking Water Sources. Environ Toxicol 23: 413-421. |
| [37] |
Burgoyne DL, Hemscheidt TK, Moore RE, et al. (2000) Biosynthesis of cylindrospermopsin. J Org Chem 65: 152-156. doi: 10.1021/jo991257m
|
| [38] |
Muenchhoff J, Siddiqui KS, Poljak A, et al. (2010) A novel prokaryotic L-arginine:glycineamidinotransferase s involved in cylindrospermopsin biosynthesis. FEBS J 277: 3844-3860. doi: 10.1111/j.1742-4658.2010.07788.x
|
| [39] |
Kellmann R, Mills T, Neilan BA (2006) Functional modeling and phylogenetic distribution of putative cylindrospermopsin biosynthesis enzymes. J Mol Evol 62: 267-280. doi: 10.1007/s00239-005-0030-6
|
| [40] | Mihali TK, Kellmann R, Muenchhoff J, et al. (2008) Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis. Appl Environ Microb74: 716-722. |
| [41] |
Lagos N, Onodera H, Zagatto HPA, et al. (1999) The first evidence of paralytic shellfish toxins in the freshwater cyanobacterium Cylindrospermopsis raciborskii, isolated from Brazil. Toxicon 37: 1359-1373. doi: 10.1016/S0041-0101(99)00080-X
|
| [42] |
Schembri MA, Neilan BA, Saint CP (2001) Identification of genes implicated in toxin production in the cyanobacterium Cylindrospermopsis raciborskii. Environ Toxicol 16: 413-421. doi: 10.1002/tox.1051
|
| [43] | Shalev-Alon G, Sukenik A, Livnah O, et al. (2002) A novel gene encoding amidinotransferase in the cylindrospermopsin producing cyanobacterium Aphanizomenon ovalisporum. FEMS Microbiol Lett 209: 87-91. |
| [44] | Neilan BA, Saker ML, Fastner J, et al. (2003). Phylogeography of the invasive cyanobacterium Cylindrospermopsis raciborskii. Mol Ecol 12: 133-140. |
| [45] |
Piccini C, AubriotL, Fabre A, et al. (2011) Genetic and eco-physiological differences of South American Cylindrospermopsis raciborskii isolates support the hypothesis of multiple ecotypes. Harmful Algae 10: 644-653. doi: 10.1016/j.hal.2011.04.016
|
| [46] |
Hoff-Risseti C, Dörr FA, Schaker PDC, et al. (2013) Cylindrospermopsin and SaxitoxinSynthetase Genes in Cylindrospermopsis raciborskii Strains from Brazilian Freshwater. PLoS ONE 8: e74238. doi: 10.1371/journal.pone.0074238
|
| [47] |
Chonudomkul D, Yongmanitchaia W, Theeragool G, et al. (2004) Morphology, genetic diversity, temperature tolerance and toxicity of Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria) strains from Thailand and Japan. FEMS Microbiol Ecol 48: 345-355. doi: 10.1016/j.femsec.2004.02.014
|
| [48] | Rasmussen JP, Giglio S, Monis PT, et al. (2008) Development and field testing of a real-time PCR assay for cylindrospermopsin-producing cyanobacteria. J Appl Microb 104: 1503-1515. |
| [49] | Stucken K, Murillo AA, Soto-Liebe K, et al. (2009) Toxicity phenotype does not correlate with phylogeny of Cylindrospermopsis raciborskii strains. Syst Appl Microbiol 32: 37-48. |
| [50] |
Froscio SM, Humpage AR, Burcham PC, et al. (2003) Cylindrospermopsin-induced protein synthesis inhibition and its dissociation from acute toxicity in mouse hepatocytes. Environ Toxicol 18: 243-251. doi: 10.1002/tox.10121
|
| [51] | Norris RLG, Seawright AA, Shaw GR, et al. (2001) Distribution of 14C Cylindrospermopsinin vivo in the Mouse. Environ Toxicol 16: 498-505. |
| [52] | Oliveira VR, CarvalhoGM, Avila MB, et al. (2012) Time-dependence of lung injury in mice acutely exposed to cylindrospermopsin. Toxicon 60: 764-772. |
| [53] |
Poniedziałek B, Rzymski P, Wiktorowicz K (2014) Toxicity of cylindrospermopsin in human lymphocytes: Proliferation, viability and cell cycle studies. Toxicol in vitro 28: 968-974. doi: 10.1016/j.tiv.2014.04.015
|
| [54] |
Poniedziałek B, Rzymski P, Wiktorowicz K (2014) Cylindrospermopsin decreases the oxidative burst capacity of human neutrophils. Toxicon 87: 113-119. doi: 10.1016/j.toxicon.2014.05.004
|
| [55] | Young FM, Micklem J, Humpage AR (2008) Effects of blue-green algal toxin cylindrospermopsin (CYN) on human granulosa cells in vitro. Reprod Toxicol 25: 374-380. |
| [56] | Gutiérrez-Praena D, Pichardo S, Jos, Á, et al. (2012) Biochemical and pathological toxic effects induced by the cyanotoxin Cylindrospermopsin on the human cell line Caco-2. Water Res 46: 1566-1575. |
| [57] | Stewart I, Wickramasinghe W, Carroll A, et al. (2012) The cylindrospermopsin analogue deoxycylindrospermopsin: Isolation, purification and acute toxicity in mice. 3rd National Cyanobacteria Workshop, Canberra, Australia, June 2012. Available from: http://www.waterra.com.au/publications/document-search/?download=697 |
| [58] | Štraser A, Filipič M, Novak M, et al. (2013) Double Strand Breaks and Cell-Cycle Arrest Induced by the Cyanobacterial Toxin Cylindrospermopsin in HepG2 Cells. Mar Drugs 11: 3077-3090. |
| [59] | Maire MA, Bazin E, Fessard V, et al. (2010) P. Morphological cell transformation of Syrian hamster embryo (SHE) cells by the cyanotoxin, Cylindrospermopsin. Toxicon 55: 1317-1322. |
| [60] | Rogers EH, Zehra RD, Gage MI, et al. (2007)The cyanobacterial toxin, cylindrospermopsin, induces fetal toxicity in the mouse after exposure late in gestation. Toxicon 49: 855-864. |
| [61] | Chernoff N, Rogers EH, Zehr RD, et al. (2011)Toxicity and recovery in the pregnant mouse after gestational exposure to the cyanobacterial toxin, cylindrospermopsin. J Appl Toxicol 31: 242-254. |
| [62] | Hayman J (1992) Beyond the Barcoo—probable human tropical cyanobacterial poisoning in outback Australia. Med J Aust 157: 794-796. |
| [63] | Carmichael WW, Azevedo SMFO, An JS, et al. (2001) Human fatalities from cyanobacteria: chemical and biological evidence for cyanotoxins. Environ Health Persp 109: 663-668. |
| [64] | Bokhari M, Carnachan RJ, Cameron NR, et al. (2007) Culture of HepG2 liver cells on three dimensional polystyrene scaffolds enhances cell structure and function during toxicological challenge. J Anat 211: 567-576. |
| [65] | Grainger SJ, Putnam AJ, 2011. Assessing the Permeability of Engineered Capillary Networks in a 3D Culture. Plos One 6: E22086. |
| [66] |
Angulo P (2002) Nonalcoholic fatty liver disease. New Engl J Med 346: 1221-1231. doi: 10.1056/NEJMra011775
|
| [67] |
Vanni E, Bugianesi E, Kotronen A, et al. (2010) From the metabolic syndrome to NAFLD or vice versa? Digest Liver Dis 42: 320-330. doi: 10.1016/j.dld.2010.01.016
|
| [68] |
Li ZZ, Berk M, McIntyre TM, et al. (2008) The Lysosomal-Mitochondrial Axis in Free Fatty Acid-Induced Hepatic Lipotoxicity. Hepatology 47: 1495-1503. doi: 10.1002/hep.22183
|
| [69] |
Ivanova S, Repnik U, Bojic L, et al. (2008) Lysosomes in apoptosis. Methods in Enzymology 442: 183-199. doi: 10.1016/S0076-6879(08)01409-2
|
| [70] | Goldman SDB, Funk RS, Rajewski RA, et al. (2009) Mechanisms of amine accumulation in, and egress from, lysosomes. Bioanalysis 1: 1445-1459. |
| [71] |
Anderson N, Borlak J (2006) Drug-induced phospholipidosis. FEBS Lett 580: 5533-5540. doi: 10.1016/j.febslet.2006.08.061
|
| [72] | Shayman JA, Abe A (2013) Drug induced phospholipidosis: An acquired lysosomal storage disorder. BiochimBiophys Acta 1831: 602-611. |
| [73] | Kornhuber J, Henkel AW, Groemer TW, et al. (2010) Lipophilic Cationic Drugs Increase the Permeability of Lysosomal Membranes in a Cell Culture System. J Cell Physiol 224: 152-164. |
| [74] | Saker ML, Thomas AD, Norton JH (1999) Cattle mortality attributed to the toxic cyanobacterium Cylindrospermopsis raciborskii in an outback region of North Queensland. Environ Toxicol 14: 179-182. |
| [75] | Shaw GR, McKenzie RA, Wickramasinghe WA, et al. (2002) Comparative Toxicity of the Cyanobacterial Toxin CylindrospermopsinBetween Mice and Cattle: Human Implications. In: Harmful Algae 2002. Xth International Conference on Harmful Algae, St. Pete Beach, Florida, U.S.A., 465-467. |
| [76] | Hamilton B, Whittle N, Shaw G, et al. (2010) Human fatality associated with Pacific ciguatoxin contaminated fish. Toxicon 56: 668-673. |
| 1. | Jingjing Wang, Hongchan Zheng, Yunfeng Jia, Dynamical analysis on a bacteria-phages model with delay and diffusion, 2021, 143, 09600779, 110597, 10.1016/j.chaos.2020.110597 | |
| 2. | WENDI WANG, DYNAMICS OF BACTERIA-PHAGE INTERACTIONS WITH IMMUNE RESPONSE IN A CHEMOSTAT, 2017, 25, 0218-3390, 697, 10.1142/S0218339017400010 | |
| 3. | Ei Ei Kyaw, Hongchan Zheng, Jingjing Wang, Htoo Kyaw Hlaing, Stability analysis and persistence of a phage therapy model, 2021, 18, 1551-0018, 5552, 10.3934/mbe.2021280 | |
| 4. | Ei Ei Kyaw, Hongchan Zheng, Jingjing Wang, Hopf bifurcation analysis of a phage therapy model, 2023, 18, 2157-5452, 87, 10.2140/camcos.2023.18.87 |

Figures(4) / Tables(3)

Ross Sadler. Towards a more complete understanding of the occurrence and toxicities of the cylindrospermopsins[J]. AIMS Environmental Science, 2015, 2(3): 827-851. doi: 10.3934/environsci.2015.3.827






 DownLoad:
DownLoad: 