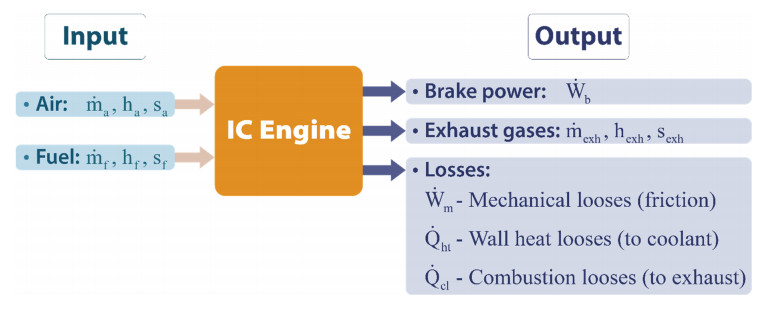
Many worldwide scientists have concentrated on using waste heat recovery technology in automotive applications because of increasingly strict fuel consumption. The thermoelectric generator (TEG) has garnered significant interest in the automobile sector as a viable waste heat recovery solution over the past several decades. A short survey of thermoelectric materials and heat exchangers for TEG systems is initially presented in this paper. To overcome the heat exchanger's current shortcomings, some previous studies designed a variety of the heat exchanger geometry of the TEG system. They suggested concentric cylindrical TEG system utilizes an annular thermoelectric module instead of the traditional square-shaped one. It uses the heat pipe's benefits to improve radial heat transmission. A comparison of the water-inside and gas-inside arrangements indicated that the water-inside concentric cylindrical TEG system provided a greater power output in our simulations to test the performance of the proposed system.
Citation: Tan Nguyen Tien, Quang Khong Vu, Vinh Nguyen Duy. Novel designs of thermoelectric generator for automotive waste heat recovery: A review[J]. AIMS Energy, 2022, 10(4): 922-942. doi: 10.3934/energy.2022042
Many worldwide scientists have concentrated on using waste heat recovery technology in automotive applications because of increasingly strict fuel consumption. The thermoelectric generator (TEG) has garnered significant interest in the automobile sector as a viable waste heat recovery solution over the past several decades. A short survey of thermoelectric materials and heat exchangers for TEG systems is initially presented in this paper. To overcome the heat exchanger's current shortcomings, some previous studies designed a variety of the heat exchanger geometry of the TEG system. They suggested concentric cylindrical TEG system utilizes an annular thermoelectric module instead of the traditional square-shaped one. It uses the heat pipe's benefits to improve radial heat transmission. A comparison of the water-inside and gas-inside arrangements indicated that the water-inside concentric cylindrical TEG system provided a greater power output in our simulations to test the performance of the proposed system.

| [1] |
Amar AB, Kouki AB, Cao H (2015) Power approaches for implantable medical devices. Sensors (Switzerland) 15: 28889–28914. https://doi.org/10.3390/s151128889 doi: 10.3390/s151128889

|
| [2] |
Jaziri N, Boughamoura A, Müller J, et al. (2020) A comprehensive review of thermoelectric generators: Technologies and common applications. Energy Rep 6: 264–287. https://doi.org/10.1016/j.egyr.2019.12.011 doi: 10.1016/j.egyr.2019.12.011
|
| [3] |
Bombarda P, Invernizzi CM, Pietra C (2010) Heat recovery from diesel engines: A thermodynamic comparison between Kalina and ORC cycles. Appl Therm Eng 30: 212–219. https://doi.org/10.1016/j.applthermaleng.2009.08.006 doi: 10.1016/j.applthermaleng.2009.08.006
|
| [4] |
Crane D, Jackson G, Holloway D (2001) Towards optimization of automotive waste heat recovery using thermoelectrics. SAE Tech Pap 1: 1–13. https://doi.org/10.4271/2001-01-1021 doi: 10.4271/2001-01-1021
|
| [5] |
Jaziri N, Boughamoura A, Müller J, et al. (2020) A comprehensive review of Thermoelectric generators: Technologies and common applications. Energy Rep 6: 264–287. https://doi.org/10.1016/j.egyr.2019.12.011 doi: 10.1016/j.egyr.2019.12.011
|
| [6] |
Karvonen M, Kapoor R, Uusitalo A, et al. (2016) Technology competition in the internal combustion engine waste heat recovery: A patent landscape analysis. J Clean Prod 112: 3735–3743. https://doi.org/10.1016/j.jclepro.2015.06.031 doi: 10.1016/j.jclepro.2015.06.031
|
| [7] |
Jia N, Cao J, Tan XY, et al. (2021) Thermoelectric materials and transport physics. Mater Today Phys 21: 100519. https://doi.org/10.1016/j.mtphys.2021.100519 doi: 10.1016/j.mtphys.2021.100519
|
| [8] |
Knecht W (2008) Diesel engine development in view of reduced emission standards. Energy 33: 264–271. https://doi.org/10.1016/j.energy.2007.10.003 doi: 10.1016/j.energy.2007.10.003
|
| [9] |
Saqr KM, Mansour MK, Musa MN (2008) Thermal design of automobile exhaust based Thermoelectric generators: Objectives and challenges. Int J Automot Technol 9: 155160. https://doi.org/10.1007/s12239-008-0020-y doi: 10.1007/s12239-008-0020-y
|
| [10] |
Zhang X, Chau KT, Chan CC (2008) Overview of Thermoelectric generation for hybrid vehicles. J Asian Electr Veh 6: 1119–1124. https://doi.org/10.4130/jaev.6.1119 doi: 10.4130/jaev.6.1119
|
| [11] |
Abedin MJ, Masjuki HH, Kalam MA, et al. (2013) Energy balance of internal combustion engines using alternative fuels. Renewable Sustainable Energy Rev 26: 20–33. https://doi.org/10.1016/j.rser.2013.05.049 doi: 10.1016/j.rser.2013.05.049
|
| [12] |
Yüksel F, Ceviz MA (2003) Thermal balance of a four stroke SI engine operating on hydrogen as a supplementary fuel. Energy 28: 1069–1080. https://doi.org/10.1016/S0360-5442(03)00090-2 doi: 10.1016/S0360-5442(03)00090-2
|
| [13] |
Durgun O, Şahin Z (2009) Theoretical investigation of heat balance in direct injection (DI) diesel engines for neat diesel fuel and gasoline fumigation. Energy Convers Manage 50: 43–51. https://doi.org/10.1016/j.enconman.2008.09.007 doi: 10.1016/j.enconman.2008.09.007
|
| [14] |
Özcan H, Söylemez MS (2006) Thermal balance of a LPG fuelled, four stroke SI engine with water addition. Energy Convers Manage 47: 570–581. https://doi.org/10.1016/j.enconman.2005.05.019 doi: 10.1016/j.enconman.2005.05.019
|
| [15] |
Mukai K, Iijima T, Miyazaki H, et al. (2005) The effects of design factors of the combustion chamber on heat balance in a gasoline engine. SAE Tech Pap 1: 1–9. https://doi.org/10.4271/2005-01-2021 doi: 10.4271/2005-01-2021
|
| [16] |
Rakopoulos CD, Giakoumis EG (2006) Second-law analyses applied to internal combustion engines operation. Prog Energy Combust Sci 32: 2–47. https://doi.org/10.1016/j.pecs.2005.10.001 doi: 10.1016/j.pecs.2005.10.001
|
| [17] |
Şahin Z, Durgun O (2007) Theoretical investigation of effects of light fuel fumigation on diesel engine performance and emissions. Energy Convers Manage 48: 1952–1964. https://doi.org/10.1016/j.enconman.2007.01.027 doi: 10.1016/j.enconman.2007.01.027
|
| [18] |
Ramadhas AS, Jayaraj S, Muraleedharan C (2006) Theoretical modeling and experimental studies on biodiesel-fueled engine. Renewable Energy 31: 1813–1826. https://doi.org/10.1016/j.renene.2005.09.011 doi: 10.1016/j.renene.2005.09.011
|
| [19] | Liu Y, Reitz RD (1998) Modeling of heat conduction within chamber walls for multidimensional internal combustion engine simulations. Int J Heat Mass Transf 41: 859–869. |
| [20] |
Lounici MS, Loubar K, Balistrou M, et al. (2011) Investigation on heat transfer evaluation for a more efficient two-zone combustion model in the case of natural gas SI engines. Appl Therm Eng 31: 319–328. https://doi.org/10.1016/j.applthermaleng.2010.09.012 doi: 10.1016/j.applthermaleng.2010.09.012
|
| [21] |
Dado B, Gelbstein Y, Dariel MP (2010) Nucleation of nanosize particles following the spinodal decomposition in the pseudo-ternary Ge0.6Sn0.1Pb0.3Te compound. Scr Mater 62: 89–92. https://doi.org/10.1016/j.scriptamat.2009.09.029 doi: 10.1016/j.scriptamat.2009.09.029
|
| [22] |
Komisarchik G, Gelbstein Y, Fuks D (2017) Solubility of Ti in thermoelectric PbTe compound. Intermetallics 89: 16–21. https://doi.org/10.1016/j.intermet.2017.05.016 doi: 10.1016/j.intermet.2017.05.016
|
| [23] | Meroz O, Gelbstein Y (2018) Thermoelectric Bi2Te3-xSex alloys for efficient thermal to electrical energy conversion. Phys Chem Chem Phys 20: 4092–4099. |
| [24] |
Sadia Y, Elegrably M, Ben-Nun O, et al. (2013) Submicron features in higher manganese silicide. J Nanomater 2013: 5–10. https://doi.org/10.1155/2013/701268 doi: 10.1155/2013/701268
|
| [25] |
Zilber T, Cohen S, Fuks D, et al. (2019) TiNiSn half-Heusler crystals grown from metallic flux for thermoelectric applications. J Alloys Comp 781: 1132–1138. https://doi.org/10.1016/j.jallcom.2018.12.165 doi: 10.1016/j.jallcom.2018.12.165
|
| [26] |
Suwardi A, Cao J, Hu L, et al. (2020) Tailoring the phase transition temperature to achieve high-performance cubic GeTe-based thermoelectrics. J Mater Chem A 8: 18880–18890. https://doi.org/10.1039/D0TA06013E doi: 10.1039/D0TA06013E
|
| [27] |
Suwardi A, Cao J, Zhao Y, et al. (2020) Achieving high thermoelectric quality factor toward high figure of merit in GeTe. Mater Today Phys 14: 100239. https://doi.org/10.1016/j.mtphys.2020.100239 doi: 10.1016/j.mtphys.2020.100239
|
| [28] |
Chen X, Dai W, Wu T, et al. (2018) Thin film thermoelectric materials: Classification, characterization, and potential for wearable applications. Coatings 8: 1–16. https://doi.org/10.3390/coatings8070244 doi: 10.3390/coatings8070244
|
| [29] |
Ren P, Liu Y, He J, et al. (2018) Recent advances in inorganic material thermoelectrics. Inorg Chem Front 5: 2380–2398. https://doi.org/10.1039/C8QI00366A doi: 10.1039/C8QI00366A
|
| [30] |
Wong-Ng W, Yan Y, Otani M, et al. (2015) High throughput screening tools for thermoelectric materials. J Electron Mater 44: 1688–1696. https://doi.org/10.1007/s11664-014-3519-1 doi: 10.1007/s11664-014-3519-1
|
| [31] |
Bhattacharya S, Madsen GKH (2015) High-throughput exploration of alloying as design strategy for thermoelectrics. Phys Rev B: Condens Matter Mater Phys 92: 1–9. https://doi.org/10.48550/arXiv.1508.03477 doi: 10.48550/arXiv.1508.03477
|
| [32] |
Gibbs ZM, Ricci F, Li G, et al. (2017) Effective mass and Fermi surface complexity factor from ab initio band structure calculations. Npj Comput Mater 3: 1–6. https://doi.org/10.1038/s41524-017-0013-3 doi: 10.1038/s41524-017-0013-3
|
| [33] |
Liu R, Chen H, Zhao K, et al. (2017) Entropy as a gene-like performance indicator promoting thermoelectric materials. Adv Mater 29: 1–7. https://doi.org/10.1002/adma.201702712 doi: 10.1002/adma.201702712
|
| [34] |
Fan Z, Wang H, Wu Y, et al. (2017) Thermoelectric performance of PbSnTeSe high-entropy alloys. Mater Res Lett 5: 187–194. https://doi.org/10.1080/21663831.2016.1244116 doi: 10.1080/21663831.2016.1244116
|
| [35] |
Fan Z, Wang H, Wu Y, et al. (2016) Thermoelectric high-entropy alloys with low lattice thermal conductivity. RSC Adv 6: 52164–52170. https://doi.org/10.1039/C5RA28088E doi: 10.1039/C5RA28088E
|
| [36] | Zhu Q, Wang S, Wang X, et al. (2021) Bottom-up engineering strategies for high-performance thermoelectric materials. Nano-Micro Letters 13: 1–38. Available from: https://link.springer.com/article/10.1007/s40820-021-00637-z. |
| [37] |
Kinsella CE, O'Shaughnessy SM, Deasy MJ, et al. (2014) Battery charging considerations in small scale electricity generation from a thermoelectric module. Appl Energy 114: 80–90. https://doi.org/10.1016/j.apenergy.2013.09.025 doi: 10.1016/j.apenergy.2013.09.025
|
| [38] |
Kawajiri K, Kishita Y, Shinohara Y (2021) Life cycle assessment of thermoelectric generators (TEGs) in an automobile application. Sustainability 13: 1–13. https://doi.org/10.3390/su132413630 doi: 10.3390/su132413630
|
| [39] |
Karri MA, Thacher EF, Helenbrook BT (2011) Exhaust energy conversion by thermoelectric generator: Two case studies. Energy Convers Manage 52: 1596–1611. https://doi.org/10.1016/j.enconman.2010.10.013 doi: 10.1016/j.enconman.2010.10.013
|
| [40] |
Liu X, Deng YD, Li Z, et al. (2015) Performance analysis of a waste heat recovery thermoelectric generation system for automotive application. Energy Convers Manage 90: 121–127. https://doi.org/10.1016/j.enconman.2014.11.015 doi: 10.1016/j.enconman.2014.11.015
|
| [41] |
Wang Y, Dai C, Wang S (2013) Theoretical analysis of a thermoelectric generator using exhaust gas of vehicles as heat source. Appl Energy 112: 1171–1180. https://doi.org/10.1016/j.apenergy.2013.01.018 doi: 10.1016/j.apenergy.2013.01.018
|
| [42] |
Liu X, Li C, Deng YD, et al. (2015) An energy-harvesting system using thermoelectric power generation for automotive application. Int J Electr Power Energy Syst 67: 510–516. https://doi.org/10.1016/j.ijepes.2014.12.045 doi: 10.1016/j.ijepes.2014.12.045
|
| [43] |
Wang P, Ji C, Tang B, et al. (2010) Effect of common rail system on vehicle engine combustion performance. Proc-2010 Int Conf Optoelectron Image Process ICOIP 2010 1: 464–467. https://doi.org/10.1109/CCIENG.2011.6007975 doi: 10.1109/CCIENG.2011.6007975
|
| [44] |
Huang K, Chang H, Qin J, et al. (2011) The exploration of HCCI combustion in the high-power direct-injection diesel engine. CCIE 2011-Proc 2011 IEEE 2nd Int Conf Comput Control Ind Eng 1: 133–135. https://doi.org/10.1109/CCIENG.2011.6007975 doi: 10.1109/CCIENG.2011.6007975
|
| [45] |
Arsie I, Cricchio A, Pianese C, et al. (2014) A comprehensive powertrain model to evaluate the benefits of electric turbo compound (ETC) in reducing CO2 emissions from small diesel passenger cars. SAE Tech Pap 1. https://doi.org/10.4271/2014-01-1650 doi: 10.4271/2014-01-1650
|
| [46] |
Hatami M, Ganji DD, Gorji-Bandpy M (2014) A review of different heat exchangers designs for increasing the diesel exhaust waste heat recovery. Renewable Sustainable Energy Rev 37: 168–181. https://doi.org/10.1016/j.rser.2014.05.004 doi: 10.1016/j.rser.2014.05.004
|
| [47] |
Thomas J (2014) Drive cycle powertrain efficiencies and trends derived from EPA vehicle dynamometer results. SAE Int J Passeng Cars-Mech Syst 7: 1374–1384. https://doi.org/10.4271/2014-01-2562 doi: 10.4271/2014-01-2562
|
| [48] |
Liming F, Wenzhi G, Hao Q, et al. (2010) Heat recovery from internal combustion engine with Rankine cycle. Asia-Pacific Power Energy Eng Conf APPEEC 1: 2–5. https://doi.org/10.1109/APPEEC.2010.5448861 doi: 10.1109/APPEEC.2010.5448861
|
| [49] |
Mingshan W, Jinli F, Chaochen M, et al. (2011) Waste heat recovery from heavy-duty diesel engine exhaust gases by medium temperature ORC system. Sci China Technol Sci 54: 2746–2753. https://doi.org/10.1007/s11431-011-4547-1 doi: 10.1007/s11431-011-4547-1
|
| [50] |
Amicabile S, Lee JI, Kum D (2015) A comprehensive design methodology of organic Rankine cycles for the waste heat recovery of automotive heavy-duty diesel engines. Appl Therm Eng 87: 574–585. https://doi.org/10.1016/j.applthermaleng.2015.04.034 doi: 10.1016/j.applthermaleng.2015.04.034
|
| [51] |
Kobayashi H, Yaji K, Yamasaki S, et al. (2021) Topology design of two-fluid heat exchange. Struct Multidiscip Optim 63: 821–834. https://doi.org/10.48550/arXiv.2005.08870 doi: 10.48550/arXiv.2005.08870
|
| [52] |
Li J, Zhou L, Pan K, et al. (1995) Evaluation of the thermodynamic process of indirect injection diesel engines by the first and second law. SAE Tech Pap: 09: 1–13. https://doi.org/10.4271/952055 doi: 10.4271/952055
|
| [53] |
Ghazikhani M, Feyz ME, Joharchi A (2010) Experimental investigation of the exhaust gas recirculation effects on irreversibility and brake specific fuel consumption of indirect injection diesel engines. Appl Therm Eng 30: 1711–1718. https://doi.org/10.1016/j.applthermaleng.2010.03.030 doi: 10.1016/j.applthermaleng.2010.03.030
|
| [54] |
Ozcan H (2010) Hydrogen enrichment effects on the second law analysis of a lean burn natural gas engine. Int J Hydrogen Energy 35: 1443–1452. https://doi.org/10.1016/j.ijhydene.2009.11.039 doi: 10.1016/j.ijhydene.2009.11.039
|
| [55] |
Rakopoulos CD, Kyritsis DC (2001) Comparative second-law analysis of internal combustion enginge operation for methane, methanol, and dodecane fuels. Energy 26: 705–722. https://doi.org/10.1016/S0360-5442(01)00027-5 doi: 10.1016/S0360-5442(01)00027-5
|
| [56] |
Saidur R, Rezaei M, Muzammil WK, et al. (2012) Technologies to recover exhaust heat from internal combustion engines. Renewable Sustainable Energy Rev 16: 5649–5659. https://doi.org/10.1016/j.rser.2012.05.018 doi: 10.1016/j.rser.2012.05.018
|
| [57] |
Duparchy A, Leduc P, Bourhis G, et al. (2009) Heat recovery for next generation of hybrid vehicles: Simulation and design of a Rankine cycle system. 24th Int Batter Hybrid Fuel Cell Electr Veh Symp Exhib 2009, EVS 24. 2: 1408–1424. https://doi.org/10.3390/wevj3030440 doi: 10.3390/wevj3030440
|
| [58] |
Conklin JC, Szybist JP (2010) A highly efficient six-stroke internal combustion engine cycle with water injection for in-cylinder exhaust heat recovery. Energy 35: 1658–1664. https://doi.org/10.1016/j.energy.2009.12.012 doi: 10.1016/j.energy.2009.12.012
|
| [59] |
Hosseini MJ, Ranjbar AA, Sedighi K, et al. (2012) A combined experimental and computational study on the melting behavior of a medium temperature phase change storage material inside shell and tube heat exchanger. Int Commun Heat Mass Transf 39: 1416–1424. https://doi.org/10.1016/j.icheatmasstransfer.2012.07.028 doi: 10.1016/j.icheatmasstransfer.2012.07.028
|
| [60] |
Jamshidi N, Farhadi M, Ganji DD, et al. (2013) Experimental analysis of heat transfer enhancement in shell and helical tube heat exchangers. Appl Therm Eng 51: 644–652. https://doi.org/10.1016/j.applthermaleng.2012.10.008 doi: 10.1016/j.applthermaleng.2012.10.008
|
| [61] |
Xu HJ, Xing ZB, Wang FQ, et al. (2019) Review on heat conduction, heat convection, thermal radiation and phase change heat transfer of nanofluids in porous media: Fundamentals and applications. Chem Eng Sci 195: 462–483. https://doi.org/10.1016/j.ces.2018.09.045 doi: 10.1016/j.ces.2018.09.045
|
| [62] |
Xu HJ, Gong L, Zhao CY, et al. (2015) Analytical considerations of local thermal non-equilibrium conditions for thermal transport in metal foams. Int J Therm Sci 95: 73–87. https://doi.org/10.1016/j.ijthermalsci.2015.04.007 doi: 10.1016/j.ijthermalsci.2015.04.007
|
| [63] |
Boomsma K, Poulikakos D (2011) Erratum: On the effective thermal conductivity of a three-dimensionally structured fluid-saturated metal foam. Int J Heat Mass Transf 54: 746–748. https://doi.org/10.1016/S0017-9310(00)00123-X doi: 10.1016/S0017-9310(00)00123-X
|
| [64] |
Brinkman HC (1952) The viscosity of concentrated suspensions and solutions. J Chem Phys 20: 571. https://doi.org/10.1063/1.1700493 doi: 10.1063/1.1700493
|
| [65] |
Buongiorno J (2006) Convective transport in nanofluids. J Heat Transfer 128: 240–250. https://doi.org/10.1115/1.2150834 doi: 10.1115/1.2150834
|
| [66] |
Calmidi VV, Mahajan RL (1999) The effective thermal conductivity of high porosity fibrous metal foams. J Heat Transfer 121: 466–471. https://doi.org/10.1115/1.2826001 doi: 10.1115/1.2826001
|
| [67] |
Das SK, Putra N, Thiesen P, et al. (2003) Temperature dependence of thermal conductivity enhancement for nanofluids. J Heat Transfer 125: 567–574. https://doi.org/10.1115/1.1571080 doi: 10.1115/1.1571080
|
| [68] |
Gao L, Zhou XF (2006) Differential effective medium theory for thermal conductivity in nanofluids. Phys Lett Sect A Gen At Solid State Phys 348: 355–360. https://doi.org/10.1016/j.physleta.2005.08.069 doi: 10.1016/j.physleta.2005.08.069
|
| [69] |
Manohara R, Harinath MA (2019) Application of Taguchi method for optimization of process parameters in drilling operation. Int J Trend Sci Res Dev 3: 1052–1057. https://doi.org/10.31142/ijtsrd24003 doi: 10.31142/ijtsrd24003
|
| [70] |
Alkidas AC, Battiston PA, Kapparos DJ (2004) Thermal studies in the exhaust system of a diesel-powered light-duty vehicle. SAE Tech Pap 1: 1–21. https://doi.org/10.4271/2004-01-0050 doi: 10.4271/2004-01-0050
|
| [71] |
Kishore RA, Kumar P, Priya S (2018) A comprehensive optimization study on Bi2Te3 -based thermoelectric generators using the Taguchi method. Sustainable Energy Fuels 2: 175–190. https://doi.org/10.1039/C7SE00437K doi: 10.1039/C7SE00437K
|
| [72] |
Kang CC, Yamauchi KA, Vlassakis J, et al. (2016) Single cell-resolution western blotting. Nat Protoc 11: 1508–1530. https://doi.org/10.1038/nprot.2016.089 doi: 10.1038/nprot.2016.089
|
| [73] |
Silaipillayarputhur K, Idem S (2013) A general matrix approach to model steady-state performance of crossflow heat exchangers. Heat Transf Eng 34: 338–348. https://doi.org/10.1080/01457632.2013.716347 doi: 10.1080/01457632.2013.716347
|
| [74] |
Silaipillayarputhur K, Al Mughanam T (2018) Performance charts for multi-pass parallel crossflow heat exchangers. Int J Mech Eng Robot Res 7: 478–482. https://doi.org/10.18178/ijmerr.7.5.478-482 doi: 10.18178/ijmerr.7.5.478-482
|
| [75] |
Romie FE (1983) Transient response of gas-to-gas crossflow heat exchangers with neither gas mixed. J Heat Transfer 105: 563–570. https://doi.org/10.1016/0017-9310(91)90249-E doi: 10.1016/0017-9310(91)90249-E
|
| [76] |
Gao T, Geer J, Sammakia B (2014) Nonuniform temperature boundary condition effects on data center cross flow heat exchanger dynamic performance. Int J Heat Mass Transf 79: 1048–1058. https://doi.org/10.1016/j.ijheatmasstransfer.2014.09.011 doi: 10.1016/j.ijheatmasstransfer.2014.09.011
|
| [77] |
Gao T, Sammakia BG, Geer FJ, et al. (2014) Dynamic analysis of cross flow heat exchangers in data centers using transient effectiveness method. IEEE Trans Components, Packag Manuf Technol 4: 1925–1935. https://doi.org/10.1109/TCPMT.2014.2369256 doi: 10.1109/TCPMT.2014.2369256
|
| [78] |
Yuan P (2003) Effect of inlet flow maldistribution on the thermal performance of a three-fluid crossflow heat exchanger. Int J Heat Mass Transf 46: 3777–3787. https://doi.org/10.1016/S0017-9310(03)00196-0 doi: 10.1016/S0017-9310(03)00196-0
|
| [79] |
Mishra M, Das PK, Sarangi S (2008) Effect of temperature and flow nonuniformity on transient behaviour of crossflow heat exchanger. Int J Heat Mass Transf 51: 2583–2592. https://doi.org/10.1016/j.ijheatmasstransfer.2007.07.054 doi: 10.1016/j.ijheatmasstransfer.2007.07.054
|
| [80] |
Guo D, Gao J, Yu YJ, et al. (2014) Design and modeling of a fluid-based micro-scale electrocaloric refrigeration system. Int J Heat Mass Transf 72: 559–564. https://doi.org/10.1016/j.ijheatmasstransfer.2014.01.043 doi: 10.1016/j.ijheatmasstransfer.2014.01.043
|
| [81] |
Jia Y, Sungtaek Ju Y (2012) A solid-state refrigerator based on the electrocaloric effect. Appl Phys Lett 100. https://doi.org/10.1063/1.4729038 doi: 10.1063/1.4729038
|
| [82] | Ossmer H, Wendler F, Gueltig M, et al. (2016) Energy-efficient miniature-scale heat pumping based on shape memory alloys. Smart Mater Struct 25: 1–13. Avaiable from: https://iopscience.iop.org/article/10.1088/0964-1726/25/8/085037. |
| [83] |
Park SIl (1996) Performance analysis of a moving-bed heat exchanger in vertical pipes. Energy 21: 911–918. https://doi.org/10.1016/0360-5442(96)00027-8 doi: 10.1016/0360-5442(96)00027-8
|
| [84] |
Rahimi A, Niksiar A (2013) A general model for moving-bed reactors with multiple chemical reactions part Ⅰ: Model formulation. Int J Miner Process 124: 58–66. https://doi.org/10.1016/j.minpro.2013.02.015 doi: 10.1016/j.minpro.2013.02.015
|
| [85] |
García-Triñanes P, Seville JPK, Ansart R, et al. (2018) Particle motion and heat transfer in an upward-flowing dense particle suspension: Application in solar receivers. Chem Eng Sci 177: 313–322. https://doi.org/10.1016/j.ces.2017.11.041 doi: 10.1016/j.ces.2017.11.041
|
| [86] |
Yusuf R, Melaaen MC, Mathiesen V (2005) Convective heat and mass transfer modeling in gas-fluidized beds. Chem Eng Technol 28: 13–24. https://doi.org/10.1002/ceat.200407014 doi: 10.1002/ceat.200407014
|
| [87] |
Zhang Y, Jiang C, Yang Z, et al. (2016) Numerical study on heat transfer enhancement in capsule-type plate heat exchangers. Appl Therm Eng 108: 1237–1242. https://doi.org/10.1016/j.applthermaleng.2016.08.033 doi: 10.1016/j.applthermaleng.2016.08.033
|
| [88] |
Song JW, Wang F, Cheng L (2012) Experimental study and analysis of a novel multi-media plate heat exchanger. Sci China Technol Sci 55: 2157–2162. https://doi.org/10.1007/s11431-012-4931-5 doi: 10.1007/s11431-012-4931-5
|
| [89] |
Villanueva HHS, de Mello PEB (2015) Heat transfer and pressure drop correlations for finned plate ceramic heat exchangers. Energy 88: 118–125. https://doi.org/10.1016/j.energy.2015.04.017 doi: 10.1016/j.energy.2015.04.017
|
| [90] |
Kim M, Baik YJ, Park SR, et al. (2010) Experimental study on corrugated crossflow air-cooled plate heat exchangers. Exp Therm Fluid Sci 34: 1265–1272. https://doi.org/10.1016/j.expthermflusci.2010.05.007 doi: 10.1016/j.expthermflusci.2010.05.007
|
| [91] |
Zhang L, Che D (2011) Influence of corrugation profile on the thermalhydraulic performance of cross-corrugated plates. Numer Heat Transf Part A Appl 59: 267–296. https://doi.org/10.1080/10407782.2011.540963 doi: 10.1080/10407782.2011.540963
|
| [92] |
Gulenoglu C, Akturk F, Aradag S, et al. (2014) Experimental comparison of performances of three different plates for gasketed plate heat exchangers. Int J Therm Sci 75: 249–256. https://doi.org/10.1016/j.ijthermalsci.2013.06.012 doi: 10.1016/j.ijthermalsci.2013.06.012
|
| [93] |
Yang J, Jacobi A, Liu W (2017) Heat transfer correlations for single-phase flow in plate heat exchangers based on experimental data. Appl Therm Eng 113: 1547–1557. https://doi.org/10.1016/j.applthermaleng.2016.10.147 doi: 10.1016/j.applthermaleng.2016.10.147
|
| [94] |
Durmuş A, Benli H, Kurtbaş I, et al. (2009) Investigation of heat transfer and pressure drop in plate heat exchangers having different surface profiles. Int J Heat Mass Transf 52: 1451–1457. https://doi.org/10.1016/j.ijheatmasstransfer.2008.07.052 doi: 10.1016/j.ijheatmasstransfer.2008.07.052
|
| [95] |
Seo JW, Kim YH, Kim D, et al. (2015) Heat transfer and pressure drop characteristics in straight microchannel of printed circuit heat exchangers. Entropy 17: 3438–3457. https://doi.org/10.3390/e17053438 doi: 10.3390/e17053438
|
| [96] | Gao T, Sammakia BG, Geer J, et al. (2014) Dynamic analysis of cross flow heat exchangers in data centers using transient effectiveness Method. IEEE Trans Components, Packag Manuf Technol 4: 1925–1935. Avaiable from: https://ieeexplore.ieee.org/document/6963402. |
| [97] |
Vintrou S, Bougeard D, Russeil S, et al. (2013) Quantitative infrared investigation of local heat transfer in a circular finned tube heat exchanger assembly. Int J Heat Fluid Flow 44: 197–207. https://doi.org/10.1016/j.ijheatfluidflow.2013.05.019 doi: 10.1016/j.ijheatfluidflow.2013.05.019
|
| [98] |
Monteiro DB, de Mello PEB (2012) Thermal performance and pressure drop in a ceramic heat exchanger evaluated using CFD simulations. Energy 45: 489–496. https://doi.org/10.1016/j.energy.2012.02.012 doi: 10.1016/j.energy.2012.02.012
|
| [99] |
Zhang C, Wang D, Han Y, et al. (2017) Experimental and numerical investigation on the exergy and entransy performance of a novel plate heat exchanger. Exp Heat Transf 30: 162–177. https://doi.org/10.1080/08916152.2016.1179358 doi: 10.1080/08916152.2016.1179358
|
| [100] |
Doo JH, Ha MY, Min JK, et al. (2012) An investigation of cross-corrugated heat exchanger primary surfaces for advanced intercooled-cycle aero engines (Part-Ⅰ: Novel geometry of primary surface). Int J Heat Mass Transf 55: 5256–5267. https://doi.org/10.1016/j.ijheatmasstransfer.2013.01.084 doi: 10.1016/j.ijheatmasstransfer.2013.01.084
|
| [101] |
Lee JM, Kwan PW, Son CM, et al. (2015) Characterizations of aerothermal performance of novel cross-corrugated plate heat exchangers for advanced cycle aero-engines. Int J Heat Mass Transf 85: 166–180. https://doi.org/10.1016/j.ijheatmasstransfer.2013.01.084 doi: 10.1016/j.ijheatmasstransfer.2013.01.084
|
| [102] |
Nilpueng K, Wongwises S (2015) Experimental study of single-phase heat transfer and pressure drop inside a plate heat exchanger with a rough surface. Exp Therm Fluid Sci 68: 268–275. https://doi.org/10.1016/j.expthermflusci.2015.04.009 doi: 10.1016/j.expthermflusci.2015.04.009
|
| [103] |
Kundu B (2015) Beneficial design of unbaffled shell-and-tube heat exchangers for attachment of longitudinal fins with trapezoidal profile. Case Stud Therm Eng 5: 104–112. https://doi.org/10.1016/j.csite.2015.03.001 doi: 10.1016/j.csite.2015.03.001
|
| [104] |
Parikshit B, Spandana KR, Krishna V, et al. (2015) A simple method to calculate shell side fluid pressure drop in a shell and tube heat exchanger. Int J Heat Mass Transf 84: 700–712. https://doi.org/10.1016/j.ijheatmasstransfer.2015.01.068 doi: 10.1016/j.ijheatmasstransfer.2015.01.068
|
| [105] |
Chang TB, Yu LY (2015) Optimal nozzle spray cone angle for triangular-pitch shell-and-tube interior spray evaporator. Int J Heat Mass Transf 85: 463–472. https://doi.org/10.1016/j.ijheatmasstransfer.2015.01.123 doi: 10.1016/j.ijheatmasstransfer.2015.01.123
|
| [106] |
Alimoradi A, Veysi F (2017) Optimal and critical values of geometrical parameters of shell and helically coiled tube heat exchangers. Case Stud Therm Eng 10: 73–78. https://doi.org/10.1016/j.csite.2017.03.003 doi: 10.1016/j.csite.2017.03.003
|
| [107] |
Raja BD, Jhala RL, Patel V (2017) Many-objective optimization of shell and tube heat exchanger. Therm Sci Eng Prog 2: 87–101. https://doi.org/10.1016/j.tsep.2017.05.003 doi: 10.1016/j.tsep.2017.05.003
|
| [108] |
Gao B, Bi Q, Nie Z, et al. (2015) Experimental study of effects of baffle helix angle on shell-side performance of shell-and-tube heat exchangers with discontinuous helical baffles. Exp Therm Fluid Sci 68: 48–57. https://doi.org/10.1016/j.expthermflusci.2015.04.011 doi: 10.1016/j.expthermflusci.2015.04.011
|
| [109] |
Salahuddin U, Bilal M, Ejaz H (2015) A review of the advancements made in helical baffles used in shell and tube heat exchangers. Int Commun Heat Mass Transf 67: 104–108. https://doi.org/10.1016/j.icheatmasstransfer.2015.07.005 doi: 10.1016/j.icheatmasstransfer.2015.07.005
|
| [110] |
Yang JF, Zeng M, Wang QW (2015) Numerical investigation on combined single shell-pass shell-and-tube heat exchanger with two-layer continuous helical baffles. Int J Heat Mass Transf 84: 103–113. https://doi.org/10.1016/j.ijheatmasstransfer.2014.12.042 doi: 10.1016/j.ijheatmasstransfer.2014.12.042
|
| [111] |
Zhou GY, Xiao J, Zhu L, et al. (2015) A Numerical Study on the Shell-Side Turbulent Heat Transfer Enhancement of Shell-and-Tube Heat Exchanger with Trefoil-Hole Baffles. Energy Procedia 75: 3174–3179. https://doi.org/10.1016/j.egypro.2015.07.656 doi: 10.1016/j.egypro.2015.07.656
|
| [112] |
Wen J, Yang H, Jian G, et al. (2016) Energy and cost optimization of shell and tube heat exchanger with helical baffles using Kriging metamodel based on MOGA. Int J Heat Mass Transf 98: 29–39. https://doi.org/10.1016/j.ijheatmasstransfer.2016.02.084 doi: 10.1016/j.ijheatmasstransfer.2016.02.084
|
| [113] |
Du BC, He YL, Wang K, et al. (2017) Convective heat transfer of molten salt in the shell-and-tube heat exchanger with segmental baffles. Int J Heat Mass Transf 113: 456–465. https://doi.org/10.1016/j.ijheatmasstransfer.2017.05.075 doi: 10.1016/j.ijheatmasstransfer.2017.05.075
|
| [114] |
Wen J, Yang H, Wang S, et al. (2017) PIV experimental investigation on shell-side flow patterns of shell and tube heat exchanger with different helical baffles. Int J Heat Mass Transf 104: 247–259. https://doi.org/10.1016/j.ijheatmasstransfer.2016.08.048 doi: 10.1016/j.ijheatmasstransfer.2016.08.048
|
| [115] |
Crane D, Lagrandeur J, Jovovic V, et al. (2013) TEG on-vehicle performance and model validation and what it means for further teg development. J Electron Mater 42: 1582–1591. https://doi.org/10.1007/s11664-012-2327-8 doi: 10.1007/s11664-012-2327-8
|
| [116] |
Mori M, Yamagami T, Sorazawa M, et al. (2011) Simulation of fuel economy effectiveness of exhaust heat recovery system using thermoelectric generator in a series hybrid. SAE Int J Mater Manuf 4: 1268–1276. https://doi.org/10.4271/2011-01-1335 doi: 10.4271/2011-01-1335
|
| [117] |
Hussain QE, Brigham DR, Maranville CW (2009) Thermoelectric exhaust heat recovery for hybrid vehicles. SAE Tech Pap 2: 1132–1142. https://doi.org/10.4271/2009-01-1327 doi: 10.4271/2009-01-1327
|
| [118] |
Espinosa N, Lazard M, Aixala L, et al. (2010) Modeling a thermoelectric generator applied to diesel automotive heat recovery. J Electron Mater 39: 1446–1455. https://doi.org/10.1007/s11664-010-1305-2 doi: 10.1007/s11664-010-1305-2
|
| [119] |
Gonçalves VM, Maia CA, Hardouin L (2012) On the solution of Max-plus linear equations with application on the control of TEGs. IFAC Proc 45: 91–97. https://doi.org/10.3182/20121003-3-MX-4033.00018 doi: 10.3182/20121003-3-MX-4033.00018
|
| [120] |
Brito FP, Goncalves LM, Martins J, et al. (2013) Influence of heat pipe operating temperature on exhaust heat thermoelectric generation. SAE Int J Passeng Cars-Mech Syst 6: 652–664. https://doi.org/10.4271/2013-01-0559 doi: 10.4271/2013-01-0559
|
| [121] | Martins J, Goncalves LM, Antunes J, et al. (2011) Thermoelectric exhaust energy recovery with temperature control through heat pipes. SAE 2011 World Congr Exhib 1: 1–16. Avaiable from: https://core.ac.uk/download/pdf/55616825.pdf. |
| [122] |
Jang JC, Chi RG, Rhi SH, et al. (2015) Heat pipe-assisted thermoelectric power generation technology for waste heat recovery. J Electron Mater 44: 2039–2047. https://doi.org/10.1007/s11664-015-3653-4 doi: 10.1007/s11664-015-3653-4
|
| [123] |
Lan S, Yang Z, Chen R, et al. (2018) A dynamic model for thermoelectric generator applied to vehicle waste heat recovery. Appl Energy 210: 327–338. https://doi.org/10.1016/j.apenergy.2017.11.004 doi: 10.1016/j.apenergy.2017.11.004
|
| [124] |
Tang ZB, Deng YD, Su CQ, et al. (2015) A research on thermoelectric generator's electrical performance under temperature mismatch conditions for automotive waste heat recovery system. Case Stud Therm Eng 5: 143–150. https://doi.org/10.1016/j.csite.2015.03.006 doi: 10.1016/j.csite.2015.03.006
|
| [125] |
Orr B, Akbarzadeh A (2017) Prospects of waste heat recovery and power generation using thermoelectric generators. Energy Procedia 110: 250–255. https://doi.org/10.1016/j.egypro.2017.03.135 doi: 10.1016/j.egypro.2017.03.135
|
| [126] |
He W, Wang S, Yue L (2017) High net power output analysis with changes in exhaust temperature in a thermoelectric generator system. Appl Energy 196: 259–267. https://doi.org/10.1016/j.apenergy.2016.12.078 doi: 10.1016/j.apenergy.2016.12.078
|
| [127] | Fankai M, Lingen Ch, Sun FR (2012) Effects of temperature dependence of thermoelectric properties on the power and efficiency of a multielement thermoelectric generator. Int J Energy Environ Eng 3: 137–150. Available from: https://www.ijee.ieefoundation.org/vol3/issue1/IJEE_14_v3n1.pdf. |
| [128] |
Gou X, Yang S, Xiao H, et al. (2013) A dynamic model for thermoelectric generator applied in waste heat recovery. Energy 52: 201–209. https://doi.org/10.1016/j.apenergy.2017.11.004 doi: 10.1016/j.apenergy.2017.11.004
|
| [129] |
Meng JH, Zhang XX, Wang XD (2014) Dynamic response characteristics of thermoelectric generator predicted by a three-dimensional heat-electricity coupled model. J Power Sources 245: 262–269. https://doi.org/10.1016/j.jpowsour.2013.06.127 doi: 10.1016/j.jpowsour.2013.06.127
|
| [130] |
Hsu CT, Huang GY, Chu HS, et al. (2011) An effective Seebeck coefficient obtained by experimental results of a thermoelectric generator module. Appl Energy 88: 5173–5179. https://doi.org/10.1016/j.apenergy.2011.07.033 doi: 10.1016/j.apenergy.2011.07.033
|

Figures(2)

Tan Nguyen Tien, Quang Khong Vu, Vinh Nguyen Duy. Novel designs of thermoelectric generator for automotive waste heat recovery: A review[J]. AIMS Energy, 2022, 10(4): 922-942. doi: 10.3934/energy.2022042







 DownLoad:
DownLoad: