Citation: Diakaridia Sangare, Ayoub Missaoui, Stéphane Bostyn, Verónica Belandria, Mario Moscosa-Santillan, Iskender Gökalp. Modeling of Agave Salmiana bagasse conversion by hydrothermal carbonization (HTC) for solid fuel combustion using surface response methodology[J]. AIMS Energy, 2020, 8(4): 538-562. doi: 10.3934/energy.2020.4.538

| [1] |
Balat M, Balat M, Kırtay E, et al. (2009) Main routes for the thermo-conversion of biomass into fuels and chemicals. Part 1: Pyrolysis systems. Energy Convers Manage 50: 3147-3157. doi: 10.1016/j.enconman.2009.08.014

|
| [2] |
McKendry P (2002) Energy production from biomass (part 1): Overview of biomass. Bioresour Technol 83: 37-46. doi: 10.1016/S0960-8524(01)00118-3
|
| [3] |
Vassilev SV, Baxter D, Andersen LK, et al. (2010) An overview of the chemical composition of biomass. Fuel 89: 913-933. doi: 10.1016/j.fuel.2009.10.022
|
| [4] | Mujtaba IM, Srinivasan R, Elbashir N (2017) The water-food-energy nexus: processes, technologies, and challenges: CRC Press. |
| [5] |
Missaoui A, Bostyn S, Belandria V, et al. (2017) Hydrothermal carbonization of dried olive pomace: Energy potential and process performances. J Anal Appl Pyrolysis 128: 281-290. doi: 10.1016/j.jaap.2017.09.022
|
| [6] |
Álvarez-Murillo A, Román S, Ledesma B, et al. (2015) Study of variables in energy densification of olive stone by hydrothermal carbonization. J Anal Appl Pyrolysis 113: 307-314. doi: 10.1016/j.jaap.2015.01.031
|
| [7] |
Benavente V, Calabuig E, and Fullana A (2015) Upgrading of moist agro-industrial wastes by hydrothermal carbonization. J Anal Appl Pyrolysis 113: 89-98. doi: 10.1016/j.jaap.2014.11.004
|
| [8] |
Hoekman SK, Broch A, Robbins C (2011) Hydrothermal carbonization (HTC) of lignocellulosic biomass. Energy Fuels 25: 1802-1810. doi: 10.1021/ef101745n
|
| [9] |
Yan W, Acharjee TC, Coronella CJ, et al. (2009) Thermal pretreatment of lignocellulosic biomass. Environ Prog Sustainable Energy: Am Inst Chem Eng 28: 435-440. doi: 10.1002/ep.10385
|
| [10] |
Tekin K, Karagöz S, Bektaş S (2014) A review of hydrothermal biomass processing. Renewable Sustainable Energy Rev 40: 673-687. doi: 10.1016/j.rser.2014.07.216
|
| [11] | Libra JA, Ro KS, Kammann C, et al. (2011) Hydrothermal carbonization of biomass residuals: A comparative review of the chemistry, processes and applications of wet and dry pyrolysis. Biofuels 2: 71-106. |
| [12] |
Hitzl M, Corma A, Pomares F, et al. (2015) The hydrothermal carbonization (HTC) plant as a decentral biorefinery for wet biomass. Catal Today 257: 154-159. doi: 10.1016/j.cattod.2014.09.024
|
| [13] |
Xiao LP, Shi ZJ, Xu F, et al. (2012) Hydrothermal carbonization of lignocellulosic biomass. Bioresour Technol 118: 619-623. doi: 10.1016/j.biortech.2012.05.060
|
| [14] |
Acharjee TC, Coronella CJ, Vasquez VR (2011) Effect of thermal pretreatment on equilibrium moisture content of lignocellulosic biomass. Bioresour Technol 102: 4849-4854. doi: 10.1016/j.biortech.2011.01.018
|
| [15] | Surup GR, Leahy JJ, Timko MT, et al. (2020) Hydrothermal carbonization of olive wastes to produce renewable, binder-free pellets for use as metallurgical reducing agents. Renewable Energy. |
| [16] | Reza MT, Nover J, Wirth B, et al. (2018) Hydrothermal carbonization of glucose in saline solution: sequestration of nutrients on carbonaceous materials. 4: 173, 2016. |
| [17] |
Kumar S, Loganathan VA, Gupta RB, et al. (2011) An assessment of U (VI) removal from groundwater using biochar produced from hydrothermal carbonization. J Environ Manage 92: 2504-2512. doi: 10.1016/j.jenvman.2011.05.013
|
| [18] |
Regmi P, Moscoso JLG, Kumar S, et al. (2012) Removal of copper and cadmium from aqueous solution using switchgrass biochar produced via hydrothermal carbonization process. J Environ Manage 109: 61-69. doi: 10.1016/j.jenvman.2012.04.047
|
| [19] |
Hu B, Wang K, Wu L, et al. (2010) Engineering carbon materials from the hydrothermal carbonization process of biomass. Adv Mater 22: 813-828. doi: 10.1002/adma.200902812
|
| [20] |
Reza MT, Rottler E, Herklotz L, et al. (2015) Hydrothermal carbonization (HTC) of wheat straw: Influence of feedwater pH prepared by acetic acid and potassium hydroxide. Bioresour Technol 182: 336-344. doi: 10.1016/j.biortech.2015.02.024
|
| [21] |
Sabio E, Álvarez-Murillo A, Román S, et al. (2016) Conversion of tomato-peel waste into solid fuel by hydrothermal carbonization: Influence of the processing variables. Waste Manage 47: 122-132. doi: 10.1016/j.wasman.2015.04.016
|
| [22] |
Chen WH, Ye SC, Sheen HK (2012) Hydrothermal carbonization of sugarcane bagasse via wet torrefaction in association with microwave heating. Bioresour Technol 118: 195-203. doi: 10.1016/j.biortech.2012.04.101
|
| [23] | Zhang L, Liu S, Wang B, et al. (2015) Effect of residence time on hydrothermal carbonization of corn cob residual. BioResources 10: 3979-3986. |
| [24] |
Chávez-Guerrero L, Hinojosa M (2010) Bagasse from the mezcal industry as an alternative renewable energy produced in arid lands. Fuel 89: 4049-4052. doi: 10.1016/j.fuel.2010.07.026
|
| [25] | Marín PC-G, Saavedra AL, Eguiarte LE, et al. (2007) En lo ancestral hay futuro: del tequila, los mezcales y otros agaves. Revista de la Universidad de Yucatán, NÚMEROS. 245: 246. |
| [26] | Gonzales B (2005) Aprovechamiento del bagazo del maguey verde (Agave salmiana) de la agroindustria del mezcal en San Luis Potosı para la producción de hongo ostra (Pleurotus ostreatus). Ph Thesis. Instituto Potosino de Investigación Cientıfica y Tecnologıca AC. México, Tesis de Maestría en Ciencias Ambientales. |
| [27] |
Michel-Cuello C, Juárez-Flores BI, Aguirre-Rivera JR, et al. (2008) Quantitative characterization of nonstructural carbohydrates of mezcal Agave (Agave salmiana Otto ex Salm-Dick). J Agric Food Chem 56: 5753-5757. doi: 10.1021/jf800158p
|
| [28] | Chávez Guerrero L (2010) Uso de bagazo de la industria mezcalera como materia prima para generar energía. Ingenierías 13: 8-16. |
| [29] |
Iñiguez-Covarrubias G, Lange SE, Rowell RM (2001) Utilization of byproducts from the tequila industry: part 1: agave bagasse as a raw material for animal feeding and fiberboard production. Bioresour Technol 77: 25-32. doi: 10.1016/S0960-8524(00)00137-1
|
| [30] |
Idarraga G, Ramos J, Zuñiga V, et al. (1999) Pulp and paper from blue agave waste from tequila production. J Agric Food Chem 47: 4450-4455. doi: 10.1021/jf990045n
|
| [31] |
Velazquez-Jimenez LH, Pavlick A, Rangel-Mendez JR (2013) Chemical characterization of raw and treated agave bagasse and its potential as adsorbent of metal cations from water. Ind Crops Prod 43: 200-206. doi: 10.1016/j.indcrop.2012.06.049
|
| [32] |
Li S, Xu S, Liu S, et al. (2004) Fast pyrolysis of biomass in free-fall reactor for hydrogen-rich gas. Fuel Process Technol 85: 1201-1211. doi: 10.1016/j.fuproc.2003.11.043
|
| [33] |
Li X-g, Ma B-g, Xu L, et al. (2006) Thermogravimetric analysis of the co-combustion of the blends with high ash coal and waste tyres. Thermochim Acta 441: 79-83. doi: 10.1016/j.tca.2005.11.044
|
| [34] |
Wang C, Wang F, Yang Q, et al. (2009) Thermogravimetric studies of the behavior of wheat straw with added coal during combustion. Biomass Bioenergy 33: 50-56. doi: 10.1016/j.biombioe.2008.04.013
|
| [35] |
Peng C, Zhai Y, Zhu Y, et al. (2016) Production of char from sewage sludge employing hydrothermal carbonization: char properties, combustion behavior and thermal characteristics. Fuel 176: 110-118. doi: 10.1016/j.fuel.2016.02.068
|
| [36] |
Guizani C, Jeguirim M, Valin S, et al. (2017) Biomass chars: The effects of pyrolysis conditions on their morphology, structure, chemical properties and reactivity. Energies 10: 796. doi: 10.3390/en10060796
|
| [37] |
Pala M, Kantarli IC, Buyukisik HB, et al. (2014) Hydrothermal carbonization and torrefaction of grape pomace: A comparative evaluation. Bioresour Technol 161: 255-262. doi: 10.1016/j.biortech.2014.03.052
|
| [38] |
Chartier A, Beaumesnil M, de Oliveira AL, et al. (2013) Optimization of the isolation and quantitation of kahweol and cafestol in green coffee oil. Talanta 117: 102-111. doi: 10.1016/j.talanta.2013.07.053
|
| [39] |
Ferreira SL, Queiroz AS, Fernandes MS, et al. (2002) Application of factorial designs and Doehlert matrix in optimization of experimental variables associated with the preconcentration and determination of vanadium and copper in seawater by inductively coupled plasma optical emission spectrometry. Spectrochim Acta Part B 57: 1939-1950. doi: 10.1016/S0584-8547(02)00160-X
|
| [40] |
Mäkelä M, Benavente V, Fullana A (2015) Hydrothermal carbonization of lignocellulosic biomass: effect of process conditions on hydrochar properties. Appl Energy 155: 576-584. doi: 10.1016/j.apenergy.2015.06.022
|
| [41] |
Linan-Montes A, De la Parra-Arciniega S, Garza-González M, et al. (2014) Characterization and thermal analysis of agave bagasse and malt spent grain. J Therm Anal Calorim 115: 751-758. doi: 10.1007/s10973-013-3321-y
|
| [42] |
Flores-Sahagun TH, Dos Santos LP, Dos Santos J, et al. (2013) Characterization of blue agave bagasse fibers of Mexico. Composites Part A: Appl Sci Manuf 45: 153-161. doi: 10.1016/j.compositesa.2012.09.001
|
| [43] |
Kambo HS, Dutta A (2015) A comparative review of biochar and hydrochar in terms of production, physico-chemical properties and applications. Renewable Sustainable Energy Rev 45: 359-378. doi: 10.1016/j.rser.2015.01.050
|
| [44] |
Liu Z, Quek A, Hoekman SK, et al. (2013) Production of solid biochar fuel from waste biomass by hydrothermal carbonization. Fuel 103: 943-949. doi: 10.1016/j.fuel.2012.07.069
|
| [45] |
Stirling RJ, Snape CE, Meredith W (2018) The impact of hydrothermal carbonisation on the char reactivity of biomass. Fuel Process Technol 177: 152-158. doi: 10.1016/j.fuproc.2018.04.023
|
| [46] |
Funke A, Ziegler F (2010) Hydrothermal carbonization of biomass: a summary and discussion of chemical mechanisms for process engineering. Biofuels, Bioprod Biorefin 4: 160-177. doi: 10.1002/bbb.198
|
| [47] |
Wiedner K, Naisse C, Rumpel C, et al. (2013) Chemical modification of biomass residues during hydrothermal carbonization-What makes the difference, temperature or feedstock? Org Geochem 54: 91-100. doi: 10.1016/j.orggeochem.2012.10.006
|
| [48] |
Lynam JG, Reza MT, Yan W, et al. (2015) Hydrothermal carbonization of various lignocellulosic biomass. Biomass Convers Biorefin 5: 173-181. doi: 10.1007/s13399-014-0137-3
|
| [49] |
Wang T, Zhai Y, Zhu Y, et al. (2018) A review of the hydrothermal carbonization of biomass waste for hydrochar formation: Process conditions, fundamentals, and physicochemical properties. Renewable Sustainable Energy Rev 90: 223-247. doi: 10.1016/j.rser.2018.03.071
|
| [50] |
Demirbas A (2004) Effects of temperature and particle size on bio-char yield from pyrolysis of agricultural residues. J Anal Appl Pyrolysis 72: 243-248. doi: 10.1016/j.jaap.2004.07.003
|
| [51] |
Collard F-X, Blin J (2014) A review on pyrolysis of biomass constituents: Mechanisms and composition of the products obtained from the conversion of cellulose, hemicelluloses and lignin. Renewable Sustainable Energy Rev 38: 594-608. doi: 10.1016/j.rser.2014.06.013
|
| [52] |
Kang S, Li X, Fan J, et al. (2012) Characterization of hydrochars produced by hydrothermal carbonization of lignin, cellulose, D-xylose, and wood meal. Ind Eng Chem Res 51: 9023-9031. doi: 10.1021/ie300565d
|
| [53] |
Magdziarz A, Wilk M, Wądrzyk M (2020) Pyrolysis of hydrochar derived from biomass-Experimental investigation. Fuel 267: 117246. doi: 10.1016/j.fuel.2020.117246
|
| [54] |
Román S, Libra J, Berge N, et al. (2018) Hydrothermal carbonization: modeling, final properties design and applications: a review. Energies 11: 216. doi: 10.3390/en11010216
|
| [55] |
Wongsiriamnuay T, Tippayawong N (2010) Non-isothermal pyrolysis characteristics of giant sensitive plants using thermogravimetric analysis. Bioresour Technol 101: 5638-5644. doi: 10.1016/j.biortech.2010.02.037
|
| [56] |
Lee Y, Park J, Ryu C, et al. (2013) Comparison of biochar properties from biomass residues produced by slow pyrolysis at 500 C. Bioresour Technol 148: 196-201. doi: 10.1016/j.biortech.2013.08.135
|
| [57] |
Wang J, Shen B, Kang D, et al. (2019) Investigate the interactions between biomass components during pyrolysis using in-situ DRIFTS and TGA. Chem Eng Sci 195: 767-776. doi: 10.1016/j.ces.2018.10.023
|
| [58] |
Basso D, Patuzzi F, Castello D, et al. (2016) Agro-industrial waste to solid biofuel through hydrothermal carbonization. Waste Manage 47: 114-121. doi: 10.1016/j.wasman.2015.05.013
|
| [59] |
Sevilla M, Fuertes AB (2009) The production of carbon materials by hydrothermal carbonization of cellulose. Carbon 47: 2281-2289. doi: 10.1016/j.carbon.2009.04.026
|
| [60] |
He C, Giannis A, Wang JY (2013) Conversion of sewage sludge to clean solid fuel using hydrothermal carbonization: hydrochar fuel characteristics and combustion behavior. Appl Energy 111: 257-266. doi: 10.1016/j.apenergy.2013.04.084
|
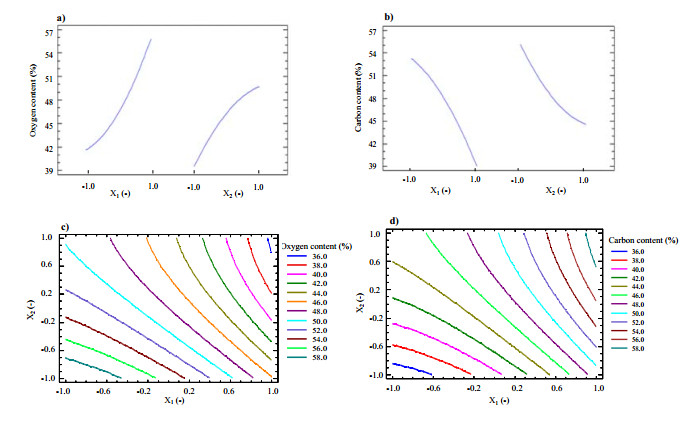
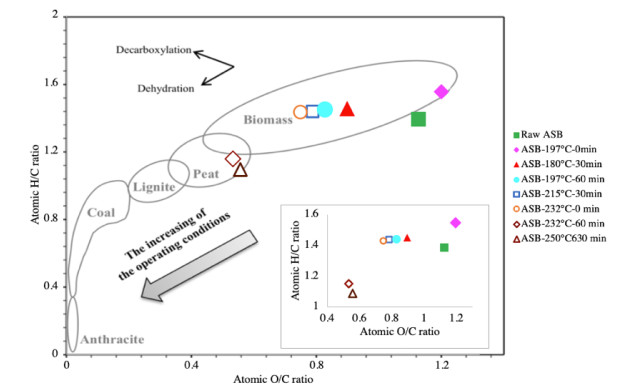
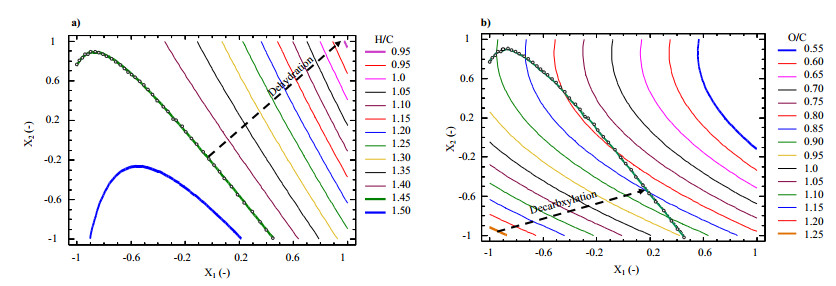
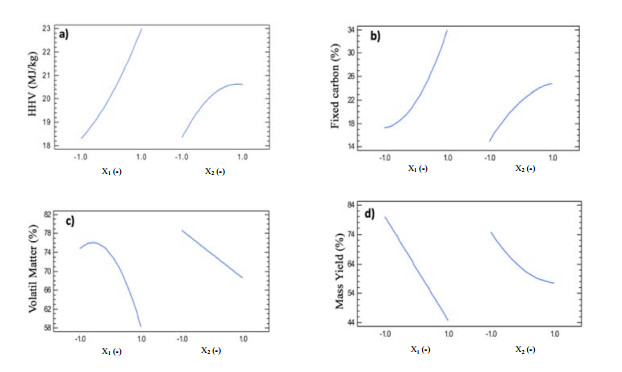
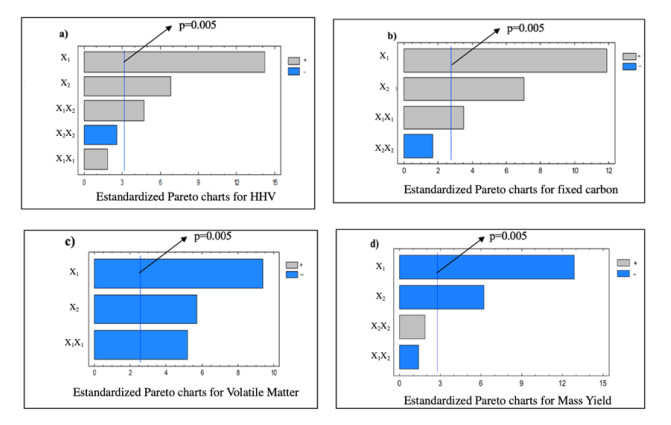
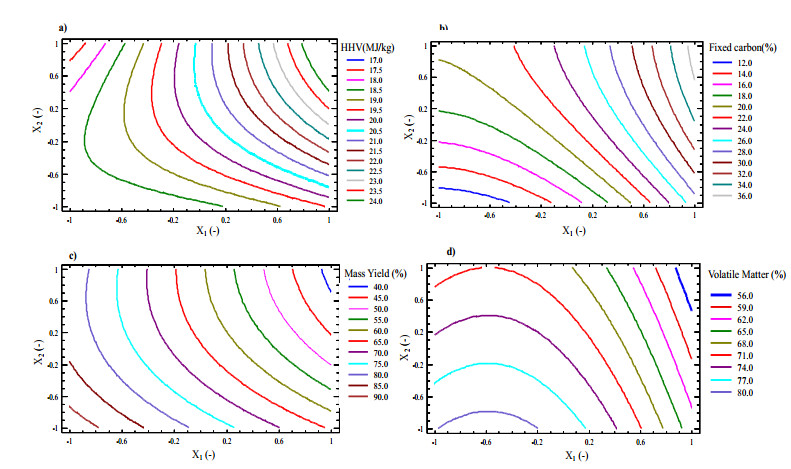
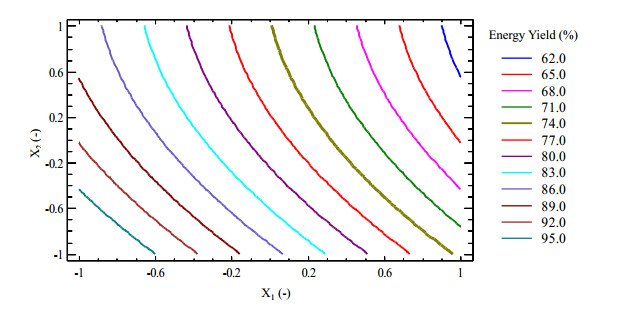
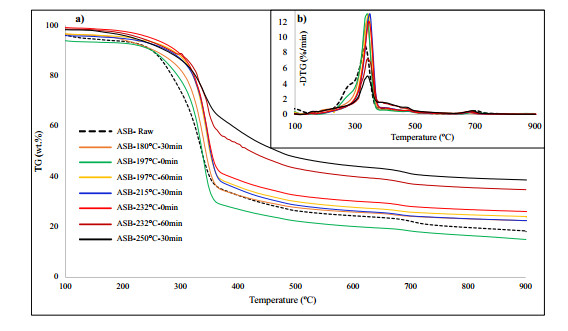
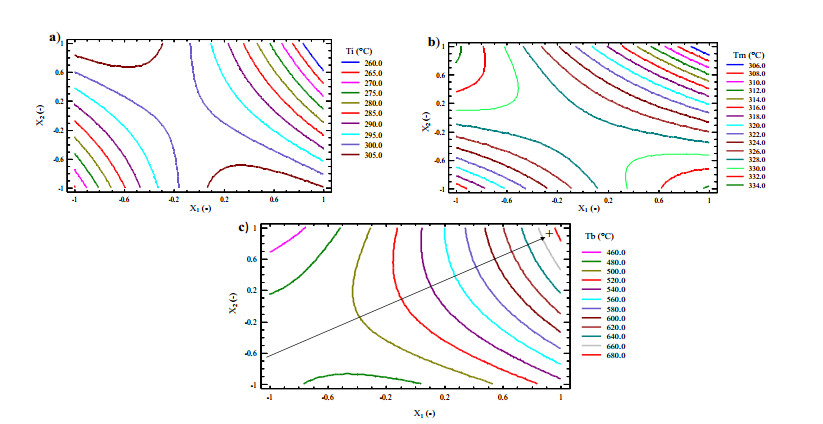
Figures(9) / Tables(5)

Diakaridia Sangare, Ayoub Missaoui, Stéphane Bostyn, Verónica Belandria, Mario Moscosa-Santillan, Iskender Gökalp. Modeling of Agave Salmiana bagasse conversion by hydrothermal carbonization (HTC) for solid fuel combustion using surface response methodology[J]. AIMS Energy, 2020, 8(4): 538-562. doi: 10.3934/energy.2020.4.538







 DownLoad:
DownLoad: