Citation: Irén Juhász Junger, Sarah Vanessa Homburg, Hubert Meissner, Thomas Grethe, Anne Schwarz Pfeiffer, Johannes Fiedler, Andreas Herrmann, Tomasz Blachowicz, Andrea Ehrmann. Influence of the pH value of anthocyanins on the electrical properties of dye-sensitized solar cells[J]. AIMS Energy, 2017, 5(2): 258-267. doi: 10.3934/energy.2017.2.258

| [1] |
O'Regan B, Grätzel M (1991) A low-cost, high-efficiency solar cell based on dye-sensitized colloidal TiO2 films. Nature 353: 737–740. doi: 10.1038/353737a0

|
| [2] |
Hagfeldt A, Boschloo G, Sun L, et al. (2010) Dye-sensitized solar cells. Chem Rev 110: 6595–6663. doi: 10.1021/cr900356p
|
| [3] | Loewenstein T, Rudolph M, Mingebach M, et al. (2010) Textile-compatible substrate electrodes with electrodeposited ZnO-a new pathway to textile-based photovoltaics. Chemphyschem 11: 783–788. |
| [4] | Almeida RH, Brito MC (2015) A review of technical options for solar charging stations in Asia and Africa. AIMS Energ 3: 428–449. |
| [5] | Macht B (2002) Degradationsprozesse in Ru(bpca)2(NCS)2-sensibilisierten Farbstoffsolarzellen auf Titandioxidbasis, Berlin, Freie Universität Berlin. |
| [6] | Hermann A, Fiedler J, Ehrmann A, et al. (2015) Strides towards textile based dye sensitized solar cells, Aachen-Dresden International Textile Conference, Aachen Aachen, Germany. |
| [7] |
Grätzel M (2003) Dye-sensitized solar cells. J Photoch Photobio C 4: 145–153. doi: 10.1016/S1389-5567(03)00026-1
|
| [8] |
Bella F, Mobarak NN, Jumaah FN, et al. (2015) From seaweeds to biopolymeric electrolytes for third generation solar cells: an intriguing approach. Electrochim Acta 151: 306–311. doi: 10.1016/j.electacta.2014.11.058
|
| [9] |
Chiappone A, Bella F, Nair JR, et al. (2014) Structure-performance correlation of nanocellulose-based polymer electrolytes for efficient quasi-solid DSSCs. Chemelectrochem 1: 1350–1358. doi: 10.1002/celc.201402051
|
| [10] |
Willgert M, Boujemaoui A, Malmström E, et al. (2016) Copper-based dye-sensitized solar cells with quasi-solid nano cellulose composite electrolytes. Rsc Adv 6: 56571–56579. doi: 10.1039/C6RA06546E
|
| [11] |
Rudhziah S, Ahmad A, Ahmad I, et al. (2015) Biopolymer electrolytes based on blend of kappa-carrageenan and cellulose derivatives for potential application in dye sensitized solar cell. Electrochim Acta 175: 162–168. doi: 10.1016/j.electacta.2015.02.153
|
| [12] | Buraidah MH, Teo LP, Yusuf SNF, et al. (2011) TiO2/Chitosan-NH4I(+I2)-BMII-based dye-sensitized solar cells with anthocyanin dyes extracted from black rice and red cabbage. Int J Photoenergy 2011: 1–11. |
| [13] | Aishah B, Nursabrina M, Noriham A, et al. (2013) Anthocyanins from hibiscus sabdariffa, melastoma malabathricum and ipomoea batatas and its color properties. Int Food Res J 20: 827–834. |
| [14] | Rakkimuthu R, Palmurugan S, Shanmugapriya A (2016) Effect of temperature, light, pH on the stability of anthocyanin pigments in cocculus hirsutus fruits. Int J Multidiscip Res Mod Educ 2: 2454–6119. |
| [15] | Oancea S, Drághici O (2013) pH and thermal stability of anthocyanin-based optimised extracts of Romanian red onion cultivars. Czech J Food Sci 31: 283–291. |
| [16] |
Ibrahim UK, Muhammad II, Salleh RM (2011) The effect of pH on color behavior of brassica oleracea anthocyanin. J Appl Sci 11: 2406–2410. doi: 10.3923/jas.2011.2406.2410
|
| [17] | Li Y, Ku SH, Chen SM, et al. (2013) Photoelectrochemistry for red cabbage extract as natural dye to develop a dye-sensitized solar cells. Int J Electrochem Sci 8: 1237–1245. |
| [18] |
Gokilamani N, Muthukumarasamy N, Thambidurai M, et al. (2013) Utilization of natural anthocyanin pigments as photosensitizers for dye-sensitized solar cells. J Sol-Gel Sci Techn 66: 212–219. doi: 10.1007/s10971-013-2994-9
|
| [19] | Alhamed M, Issa AS, Doubal AW (2012) Studying of natural dyes properties as photo-sensitizer for dye sensitized solar cells (DSSC). J Electron Dev 16: 1370–1383. |
| [20] |
Kumara NTRN, Ekanayake P, Lim A, et al. (2013) Study of the enhancement of cell performance of dye sensitized solar cells sensitized with nephelium lappaceum (F: Sapindaceae). J Sol Energ Eng 135: 031014–031018. doi: 10.1115/1.4023877
|
| [21] |
Shahzad N, Pugliese D, Lamberti A, et al. (2013) Monitoring the dye impregnation time of nanostructured photoanodes for dye sensitized solar cells. J Phys Conf Ser 439: 012012–012023. doi: 10.1088/1742-6596/439/1/012012
|
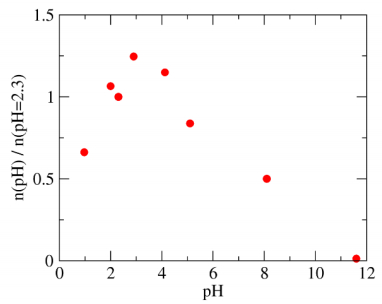
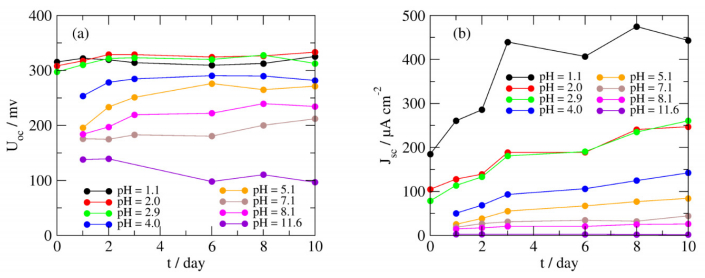
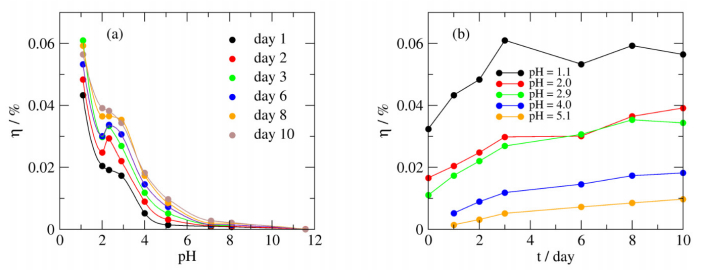
Figures(7)

Irén Juhász Junger, Sarah Vanessa Homburg, Hubert Meissner, Thomas Grethe, Anne Schwarz Pfeiffer, Johannes Fiedler, Andreas Herrmann, Tomasz Blachowicz, Andrea Ehrmann. Influence of the pH value of anthocyanins on the electrical properties of dye-sensitized solar cells[J]. AIMS Energy, 2017, 5(2): 258-267. doi: 10.3934/energy.2017.2.258







 DownLoad:
DownLoad: