Citation: Hamid Shahandeh, Frank M. Hons, Joseph O. Storlien, Jason P. Wight. Long-term bioenergy sorghum harvest strategy and soil quality[J]. AIMS Energy, 2016, 4(4): 633-657. doi: 10.3934/energy.2016.4.633
| [1] | Hamid Shahandeh, Frank M. Hons, Jason P. Wight, Joseph O. Storlien . Harvest strategy and N fertilizer effects on bioenergy sorghum production. AIMS Energy, 2015, 3(3): 377-400. doi: 10.3934/energy.2015.3.377 |
| [2] | Kevin R Caffrey, Mari S Chinn, Matthew W Veal . Biomass supply chain management in North Carolina (part 1): predictive model for cropland conversion to biomass feedstocks. AIMS Energy, 2016, 4(2): 256-279. doi: 10.3934/energy.2016.2.256 |
| [3] | Robinson J. Tanyi, Muyiwa S Adaramola . Bioenergy potential of agricultural crop residues and municipal solid waste in Cameroon. AIMS Energy, 2023, 11(1): 31-46. doi: 10.3934/energy.2023002 |
| [4] | Kevin Caffrey, Mari Chinn, Matthew Veal, Michael Kay . Biomass supply chain management in North Carolina (part 2): biomass feedstock logistical optimization. AIMS Energy, 2016, 4(2): 280-299. doi: 10.3934/energy.2016.2.280 |
| [5] | Romain Akpahou, Marshet M. Admas, Muyiwa S Adaramola . Evaluation of a bioenergy resource of agricultural residues and municipal solid wastes in Benin. AIMS Energy, 2024, 12(1): 167-189. doi: 10.3934/energy.2024008 |
| [6] | Eric Danso-Boateng, Osei-Wusu Achaw . Bioenergy and biofuel production from biomass using thermochemical conversions technologies—a review. AIMS Energy, 2022, 10(4): 585-647. doi: 10.3934/energy.2022030 |
| [7] | Patrick Moriarty, Damon Honnery . Review: Assessing the climate mitigation potential of biomass. AIMS Energy, 2017, 5(1): 20-38. doi: 10.3934/energy.2017.1.20 |
| [8] | Thananat Lungkadee, Tossapon Katongtung, Pich-ramon Pokkanta, Tossaporn Onsree, Chawannat Jaroenkhasemmeesuk, Nakorn Tippayawong . Current technology landscape of biochar in carbon capture and storage research via bibliometric analysis. AIMS Energy, 2024, 12(1): 277-303. doi: 10.3934/energy.2024014 |
| [9] | Noelia M. Elía, Sue E Nokes, Michael D. Flythe . Switchgrass (Panicum virgatum) fermentation by Clostridium thermocellum and Clostridium saccharoperbutylacetonicum sequential culture in a continuous flow reactor. AIMS Energy, 2016, 4(1): 95-103. doi: 10.3934/energy.2016.1.95 |
| [10] | Patrick Moriarty, Damon Honnery . The Earth We are Creating. AIMS Energy, 2014, 2(2): 158-171. doi: 10.3934/energy.2014.2.158 |
Bioenergy sorghum [Sorghum bicolor (L.) Moench.] is among the proposed agricultural crops to assist the U.S. in developing renewable bioenergy and reducing greenhouse gas (GHG) emissions [1,2,3]. Bioenergy sorghum is a novel second generation crop that does not compete directly with food crops, possesses high biomass yield potential, is drought tolerant, and has established production systems and a tractable breeding improvement program [1,4]. Texas and the Southern Great Plains have been proposed as the most reliable areas for bioenergy sorghum production [5]. However, removal of biomass for energy may potentially have adverse effects on soil productivity and on soil and environmental quality [6,7]. Therefore, it is critical to examine the consequences of removing produced biomass on SOC and other soil properties before biofuel production expands [8].
Muth et al. [9] conducted a comprehensive study of crop residue removal that ranged from 22% to 83% for bioenergy production at the soil-type level across the U.S. Their analysis showed that over both the short- and long-term, 150 and 208 million metric tons of agricultural residues could be sustainably removed for bioenergy production. Continued removal of crop residue, however, can negatively impact soil properties including SOC, TSN and can also decrease aggregate stability, soil microbial biomass, and particulate organic matter, while increasing soil bulk density [10]. According to the USDA NRCS [11], monitoring of SOC should be conducted to ensure that soil quality is not sacrificed in the name of renewable bioenergy, and should not jeopardize soil quality or SOC in long-term cropping [8,12,13,14]. The removal of crop residues for bioenergy feedstock, or other purposes, should be done with caution to avoid loss of soil organic matter (SOM) [15].
Soil organic C is the most reported indicator of soil quality and is an important attribute for sustainable crop production [16,17] in that it impacts many soil physical, chemical and biological properties [16,18]. Follett et al. [19] stated that sustainability of bioenergy crop production depends on SOC sequestration. However, one direct source of the SOC pool is crop residues [20] and removal of crop residues can deplete SOC pools [6]. Residue contributions to the SOC pool are influenced by cropping systems, tillage methods and nutrient applications. For example, N fertilizer or crop rotation has shown to be important for residue production and has increased SOC sequestration under different cropping systems [19,21,22,23,24].
Crop residues also contain substantial amounts of plant-essential nutrients [25] that are removed when biomass is harvested. Therefore, in order to maintain the productivity of many soils, it will be essential to recycle mineral nutrients from harvested biomass or animal manures or to directly add nutrients to soil as chemical fertilizers. Somerville et al. [26] expressed concern that removal of even half the stover produced would exacerbate SOC loss and would require additional inputs of fertilizers to replace lost nutrients. Reijnders [14] suggested that in order to sustain fertility of cropland used for bioenergy, nutrients in treated residues following biofuel production should be returned to soil. Findings of Kirkby et al. [27] indicated that inadequate nutrient availability limits SOC sequestration in arable soils and suggested that optimum C sequestration requires additional nutrients beyond that required for crop production alone.
However, little is known about the effect of bioenergy crop residue removal on SOC storage, with major uncertainty regarding the impacts of cropping intensification on SOC loss and the quantities of crop residue that can be sustainably harvested without jeopardizing SOC stocks [8,12,13,28]. Studies on sustainable corn stover removal thresholds indicated that perhaps 30-50% of harvestable biomass could be removed while maintaining SOC [15,29,30]. Anderson-Teixeira et al. [31] estimated that corn stover removal rates of 25 and 100% would reduce the SOC pool by about 3 and 8 Mg ha−1 yr−1 in the 0 to 30 cm soil depth. Johnson et al. [32] empirically estimated the minimum corn residue return rate required to sustain SOC levels at numerous locations. Based on duration and a relationship between the rate of residue returned and changes in measureable SOC, a minimum return rate of 3.9 ± 2.18 Mg corn stover ha−1 yr−1 was identified. Wilhelm et al. [7,33] and Johnson et al. [34] suggested that a minimum of about 5.3 Mg ha−1 of corn residue was necessary to maintain SOC in no till systems, while Halvorson et al. [35] suggested 6.8 to 7.6 Mg ha−1 yr−1 under limited irrigation. Depending on cropping system, crop species, tillage management, and climate, estimates vary from 0.8 to 14 Mg residue ha−1 for maintaining SOC [7,34,36,37].
Information on harvest thresholds and soil quality for energy sorghum is scarce or has been based on modeled assumptions from short-term research [12,13,38]. Meki et al. [13], using models to evaluate several biomass sorghum removal rates and tillage systems, predicted that 75% biomass removal was sustainable for no till bioenergy sorghum production, resulting in annual harvestable biomass yield of 18.0 ± 0.9, biomass residue return of 6 ± 0.3, and a root biomass of 7.2 ± 0.4 Mg ha−1.
The impacts of crop residue removal on SOC has been reviewed for common agricultural production systems [6,39] and mitigation strategies have been proposed, but management strategies that might be used to counteract SOC losses from residue removal for bioenergy crops have not been widely discussed. Sainju et al. [38] used cover crops to increase soil C and N storage under bioenergy sorghum production, and suggested these increases might have been most associated with contributions from roots. In fact, in order to properly quantify the amount of crop C incorporated into SOC, belowground as well as aboveground biomass produced should be determined. Mazzilli et al. [40] estimated the contribution of aboveground and belowground biomass to the formation of SOC in four agricultural treatments by measuring changes in 13C natural abundance in particulate organic matter (CPOM) associated with manipulations of C3 and C4 biomass. After two growing seasons, CPOM was primarily derived from belowground C inputs, even though they represented only ~10% of the total plant C inputs from residues. Belowground biomass contributed from 60% to almost 80% of the total new C present in the CPOM in the top 10 cm of soil. Their results indicated that roots may play a disproportionately important role in the CPOM budget in soils. Simulation modeling estimates of Meki et al. [12] predicted that, on average, root biomass contributed 24%, 37%, 54%, and 95%, respectively, of total sorghum biomass inputs when aboveground biomass was removed at 0%, 50%, 75%, and 100%. Wilhelm et al. [7] reported that roots contributed more than half of the soil C input, while Flesa et al. [41] concluded that 37% of root biomass was incorporated into SOC. Wilts et al. [42] studied root- derived C in a long-term study of corn stover harvest, and concluded that total C from the unharvestable portion of the crop (mainly roots) was 1.7 to 1.8 times more than from aboveground sources whether or not fertilizer N was added. Liebig et al. [43] stated that increased SOC under switchgrass was likely caused by belowground C input from both root biomass and rhizodeposition. Schmidt et al. [44] suggested that SOM (especially in deeper soil layers) originates from root exudates and microbial products rather than incorporation of above ground biomass into deeper soils, and is further supported by growing deep rooted crops like switchgrass and alfalfa to maintain SOC deeper in soils [43].
Most research on SOC changes in agricultural production systems has focused on the 0 to 30-cm depth [7,36]. A main objective of the current research study was to determine effects of rotation, sorghum biomass return, and N fertilization on the C economy and soil quality indicators of bioenergy sorghum production systems to 90-cm depth.
The experimental site was located at the Texas A&M AgriLife Research Farm, approximately 8 km southwest of College Station, TX (30°3'15ʺ N lat; 96°25ʹ37ʺ W long), which is situated within the Brazos River floodplain in south-central Texas [24]. The soil used was a calcareous (pH 8.2) Weswood silty clay loam (fine-silty, mixed, superactive, thermic Udifluventic Haplustept), with particle size distribution of 100, 560, and 340 g kg−1 of sand, silt, and clay, respectively, in the top 15 cm of soil. Climate at the site is classified as humid subtropical, with mean annual precipitation of 1017 mm and mean annual temperature of 20 °C. Before the start of the bioenergy sorghum study in 2008, the field was in cotton [Gossypium hirsutum (L.)] in 2007 and had been rotated biannually with corn under conventional disk tillage for the previous ten years. Soil nutrient properties at the initiation of the study and detailed field management practices performed were reported by Wight et al. [45]. The study soil initially rated very low for extractable NO3-N, moderate for P, and high for K, Ca and Mg [45].
The experimental design was a randomized complete block with either continuous bioenergy sorghum or sorghum rotated biannually with corn, two N rates (0 and 280 kg N ha−1 for sorghum; 0 and 150 kg N ha−1 for corn), three levels of sorghum residue (biomass) return (0, 25% and 50%; 100% of corn stover was returned), and four replications. An additional nutrient return treatment received 280 kg N ha−1 and complete sorghum residue removal at harvest. Biomass yield from this treatment was determined each year, and was subsequently analyzed for total concentrations of P, K, Ca, Mg, Fe, Mn, Zn and Cu as described below. The amounts of these elements removed annually were subsequently returned to these treatment plots in inorganic form prior to the next growing season. The study utilized four row plots (1.02-m row centers) measuring 9.14 m long and 4.08 m wide. The bioenergy sorghum cultivar used, “4Ever Green, ” was a photoperiod sensitive, high-yielding hybrid forage sorghum (Walter Moss Seed Co., Waco, TX), while the corn hybrids were DKC64-69 and DKC68-05 (DeKalb Brand, Monsanto Co., St. Louis, MO). Corn grain and aboveground biomass were hand-harvested for yield at maturity (July 2nd-28th) from a randomly selected 3-m segment of the middle two rows from each plot. Stover from remaining non-sampled portions of the corn plots was not incorporated into soil until at the later sorghum harvesting date. All corn residue was ultimately returned to each plot, regardless of sorghum residue return treatment. Field management practices were performed as described by Wight et al. [45]. Limited furrow irrigation was used to prevent severe water stress. Treatments receiving N fertilization were side-dressed with subsurface banded granular urea at approximately the four-leaf stage for sorghum and six-leaf stage for corn. Sorghum harvest was performed with a silage harvester with an attached weigh-bucket and scale, and occurred between August 5th and September 13th for all years, except for October and November harvests in 2008 and 2009 [45]. Biomass yield was estimated from the entire length of the middle two rows of each plot, and a random grab sample of chopped residue was captured for determining moisture and nutrient composition. The harvester chopped sorghum biomass into approximately 2 cm by 2 cm pieces. To physically simulate harvest strategies that would return 25% or 50% of the sorghum crop residue, biomass harvested from the middle one or two rows was evenly distributed across the area of the entire plot. All treatments were imposed in 2008 and continued annually through 2014.
Baseline composite soil samples for each plot were initially collected in March 2008 prior to planting or treatment imposition using three, 3.8-cm diameter soil cores at 0-5, 5-15, 15-30, 30-60, and 60-90 cm. Similar samples were taken for each plot prior to each growing season between March 7th and April 5th from 2009-2015. Samples were oven-dried at 60 °C for 7 days, ground using a flail grinder and sieved to pass through a 1.75- mm mesh screen. Residual soil inorganic nitrate-N and extractable soil nutrients were measured and a subsample of each composite soil sample was also finely ground in a ring and puck mill and analyzed for organic and inorganic C and total N [45].
Soil C and N content in each depth increment was calculated by multiplying soil C and N concentration by soil bulk density which was estimated from measured SOC [46]. The bulk density of soil for each depth increment initially was determined by dividing the weight of oven dried soil by the volume of the core. The core method gave reasonable bulk density values in the top 30 cm of soil with narrow depth increments [39,47] but beyond 30 cm with larger increments of 30 to 60 and 60 to 90 cm, bulk density measurements became unreasonably high. Therefore, the Adams [46] method was used to determine bulk density for all depth increments.
Sorghum was planted at a seeding density of 160, 000 seed ha−1 between March 19th and April 17th during the seven years of study using a four-row planter (Case IH). The biomass was harvested using a New Holland model 707U forage chopper (New Holland, PA, USA) that resulted in 18.65 m2 of field area for determination of each plot yield. Approximately 600 g of chopped plant material was taken as a subsample and immediately weighed. Subsamples then were oven dried at 60 °C until stable weights were attained. Dried plant samples were initially ground to pass a 1-mm sieve and then powdered in a ring and puck mill prior to total elemental analysis for C and N by combustion [45,48]. Biomass C and N contents were calculated by multiplying dry biomass yields by biomass C and N concentrations. The average C concentration of sorghum biomass was approximately 42%, but ranged from 41 to 44% (data not shown). Inorganic constituents of biomass from the nutrient return treatment were determined following nitric acid digestion [45].
The effects of N fertilization, biomass return, and their interactions on biomass yield and plant and soil properties were tested using PROC Mixed in SAS 9.2 (SAS Institute (2010).All differences discussed were significant at the P ≤ 0.05 probability level or less. A least significant difference (LSD) was calculated if overall treatment significance (P < 0.05) was established using ANOVA.
Nitrogen fertilization significantly increased biomass and biomass C yields of both continuous and rotated sorghum, but the rate of increase was influenced by the amount of residue (biomass) returned to soil for continuous sorghum (Figure 1). Biomass and biomass C yields of rotated sorghum were higher than those of continuous sorghum regardless of N fertilization or residue return. Residue return had no significant effect on biomass or biomass C yield of rotated sorghum whether or not sorghum was fertilized. In contrast, the effect of residue return on biomass and biomass C yield was significant when continuous bioenergy sorghum was fertilized (Figure 1). Biomass and biomass C yields averaged across residue return rates for rotated sorghum were 22.9 and 9.6 Mg ha-1 and 19.1 and 8.1 ha−1, respectively, with and without N fertilization. Corresponding biomass and biomass C yields for continuous sorghum across residue return were 17.8 and 7.5 and 11.5 and 4.8 Mg ha−1 with and without N fertilization, or approximately 55 and 20% yield increases for continuous and rotated sorghum from fertilization. Wight et al. [45] also reported greater biomass yield increases from N fertilization of continuous compared with rotated biomass sorghum. Hons et al. [49] reported both higher cotton lint yields and N use efficiency for cotton following corn compared to continuous cotton on a soil similar to that used in our study. Higher biomass yields of rotatedsorghum might potentially be related to nutrient cycling from corn stover or from the often reported beneficial effects of rotation through improved soil physical condition [50], a reduction in both pathogenic [51] and non-pathogenic deleterious microorganisms [52], the proliferation of beneficial soil microorganisms, including mycorrhizae [53], and the breaking of insect and disease cycles. However, not all mechanisms appear to be important in all cropping systems [54]. The allelopathic influence of sorghum residues on crops has also been observed in both monoculture and multiple cropping systems [55] and may be another reason for lower yields with continuous sorghum.
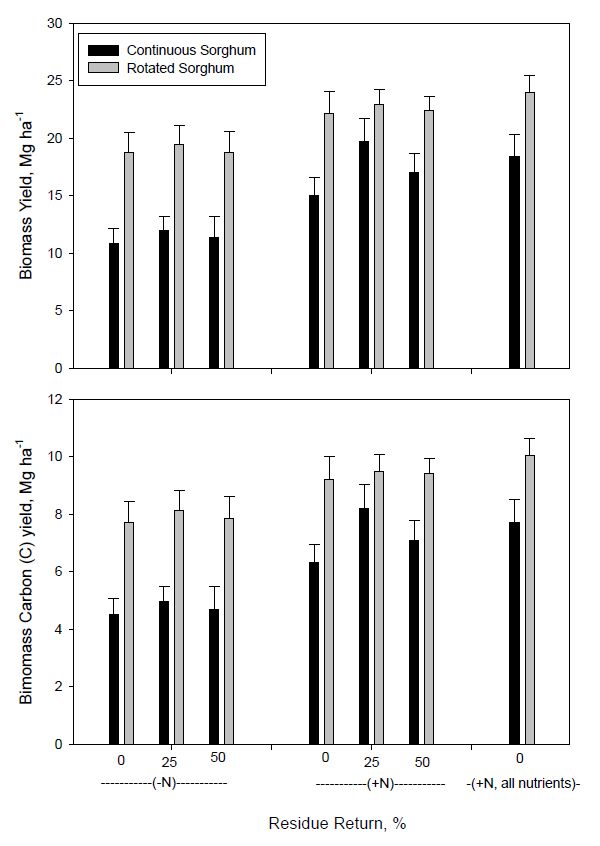 Figure 1. Average biomass and biomass C yields for continuous and rotated sorghum for six and three years, respectively, during 2009 to 2014 as influenced by N fertilization and biomass return rate. All nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.
Figure 1. Average biomass and biomass C yields for continuous and rotated sorghum for six and three years, respectively, during 2009 to 2014 as influenced by N fertilization and biomass return rate. All nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.Nitrogen fertilization has consistently been reported to increase bioenergy sorghum yields [13,24,45,56,57]. Addition of residue without added N was not sufficient to support maximum long-term bioenegy sorghum production (Figure 1). Shahandeh et al. [24] reported that even with 50% biomass return, sorghum biomass yield was reduced by more than 35% after 6 years when no N was applied. Meki et al. [13] used a simulation model to show that long-term sorghum biomass removal without N application would result in reduced total biomass yields due to soil N depletion and eventual soil nutrient mining. The nutrient return treatment in our study resulted in either the greatest or statistically equivalent biomass yield for both rotated and continuous bioenergy sorghum (Figure 1). The greatest yieldincrease from residue return was observed with 25% return and N addition with continuous sorghum, having biomass and biomass C yields statistically similar to that for rotated sorghum receiving this treatment. Continuous sorghum had lower yields than rotated for all other treatments.
The amounts of harvested sorghum biomass returned to soil and corresponding C and N contents and C:N ratios are shown in Table 1. Obviously, more sorghum biomass was returned to soil in 6 years of continuous sorghum cropping than in 3 years of rotated sorghum, especially with N fertilization. For example, at 25% return in continuous sorghum, the cumulative input of total biomass returned was increased by about 64% (21.0 to 34.5 Mg ha−1) over 6 years by N fertilization, while at 50% return, biomass returned increased by 51% (39.7 to 59.9 Mg ha−1). Cumulative residue returned over 3 years with rotated sorghum and 25% residue return was increased by 14% (14.6 to 16.7 Mg ha−1) with N fertilization, while the corresponding increase was 19% (28.2 to 33.6 Mg ha−1) with 50% residue return. Model simulations conducted by Meki et al. [13] suggested the minimum sorghum biomass return needed to maintain and/or increase SOC, and hence protect soil quality, ranged between 6.2 and 14.8 Mg ha−1 annually, indicating that the biomass removal threshold is subject to many site specific factors. These authors predicted a permissible biomass removal range of 50-75% for bioenergy sorghum, which compared reasonably well with a suggested corn stover removal of 67 to 74%, returning a minimum of 5.7 Mg stover ha−1yr−1 [34]. The annual sorghum residue returned at the 50% rate in our study varied from 4.7 Mg ha−1 yr−1 for rotated sorghum without N fertilization to 10.0 Mg ha−1 yr−1 for continuous sorghum with N fertilization, respectively (Table 1).
| Fertilization | Residue Return | Continuous Sorghum | Rotated Sorghum | ||||||
| Biomass | C | N | C:N | Biomass | C | N | C:N | ||
| % | ---Mg ha-1 | kg ha-1 | ---Mg ha-1 | kg ha-1 | |||||
| Total Return | |||||||||
| -N | 25 | 21.0 | 8.7 | 162 | 54 | 14.6 | 6.1 | 145 | 42 |
| -N | 50 | 39.7 | 16.4 | 288 | 57 | 28.2 | 11.8 | 284 | 42 |
| +N | 25 | 34.5 | 14.4 | 318 | 45 | 16.7 | 6.9 | 217 | 32 |
| +N | 50 | 59.9 | 24.8 | 530 | 47 | 33.6 | 14.1 | 423 | 33 |
| Annual Return † | |||||||||
| -N | 25 | 3.5 | 1.5 | 27 | 55 | 2.4 | 1.0 | 24 | 42 |
| -N | 50 | 6.6 | 2.7 | 48 | 56 | 4.7 | 2.0 | 47 | 41 |
| +N | 25 | 5.8 | 2.4 | 53 | 45 | 2.8 | 1.2 | 36 | 32 |
| +N | 50 | 10.0 | 4.1 | 88 | 47 | 5.6 | 2.4 | 71 | 33 |
| † Average of 6 years. | |||||||||
 DownLoad: CSV
DownLoad: CSVGreater amounts of harvested residue returned with N fertilization also resulted in larger quantities of C and N and lower C:N ratios in returned residue for both continuous and rotated bioenergy sorghum (Table 1). Trends for quantities of biomass C returned tended to follow those for biomass yield since biomass C concentrations were similar with and without N fertilization. Biomass N returned with N fertilization tended to be greater on a relative basis, however, because of the greater proportional increase in biomass N concentration with fertilization. The average C:N ratio of both continuous and rotated sorghum biomass was about 10 units higher within residue return rate without than with N fertilization (Table 1). Average biomass C:N for continuous sorghum ranged from 12 to 15 units higher compared to corresponding treatments with rotated sorghum. Cadoux et al. [58] also reported that sorghum biomass C:N was lowered by N application, while Halvorson and Stewart [59] observed that corn stover C:N declined with increasing N addition rate. Bioenergy sorghum biomass also had lower C:N ratios than for many cereal crops [56].
The C:N ratio of crop residue is considered to be an important parameter controlling the C sequestration potential of various crops, and along with stover/residue yield, has been used to help estimate the C sequestration potential of cropping systems [56,60]. Biomass with wider C:N ratios may take longer to decompose, thereby potentially increasing SOC storage time and slowing the release of CO2 to the atmosphere. Decreased biomass C:N ratios with N fertilization might result in faster decomposition and therefore a larger amount of returned residue required to maintain SOC. The amount of crop residue C converted to stable SOC depends on the amount and quality of the residue and cropping management [34,61,62].
Soil organic C concentrations in 2015 under continuous sorghum receiving residue return and N fertilization were increased at all soil depths, except 15-30 cm, compared to initial samples (Table 2). Residue return withoutadded N also significantly increased SOC at 0-5 cm, although increases were smaller compared to corresponding treatments receiving N, and were not observed at deeper depths. Increasing residue return from 25 to 50% tended to increase SOC in the near surface regardless of N addition, but generally not with depth. The largest relative SOC increases were observed under continuous sorghum receiving N fertilization and 25% residue return at 30-60 and 60-90 cm depths with 49% (4.9 to 7.3 g kg−1), and 81% (3.7 to 6.7 g kg−1) increases, respectively. Soil organic C concentrations with N fertilization under continuous sorghum averaged across residue return rates increased by 20, 10, 13, 31 and 44%, respectively, at 0-5, 5-15, 15-30, 30-60, and 60-90 cm. The nutrient return treatment also increased SOC at 0-5, 5-15, and 30-60 cm compared with initial values. After three years of bioenergy sorghum without added N in the corn/sorghum rotation, SOC concentrations wereeither similar to initial values, or slightly decreased, even with residue return (Table 2). Nitrogen fertilization only increased SOC concentrations at 0-5 cm under this cropping sequence.
| Fertilization | Residue Return | Soil Depth (cm) | ||||
| 0–5 | 5–15 | 15–30 | 30–60 | 60–90 | ||
| % | Continuous Sorghum, SOC, g kg-1 | |||||
| -N | 0 | 9.8 (9.3†) | 8.3 (8.0) | 7.1 (6.5) | 4.5 (4.9) | 3.6(3.7) |
| -N | 25 | 11.6* | 8.5 | 7.3 | 4.9 | 3.7 |
| -N | 50 | 12.0* | 8.9 | 6.2 | 3.9 | 4.0 |
| +N | 0 | 10.1* | 8.1 | 7.4 | 5.7* | 3.7 |
| +N | 25 | 12.1* | 9.0* | 7.6 | 7.3* | 6.7* |
| +N | 50 | 12.8* | 9.4* | 7.1 | 6.2* | 5.6* |
| +N, nutrients | 0 | 10.2* | 9.1* | 7.6 | 6.4* | 4.6 |
| Rotated Sorghum, SOC, g kg-1 | ||||||
| -N | 0 | 9.6 (9.3†) | 7.5 (8.0) | 5.6* (6.5) | 3.9 (4.9) | 3.2(3.7) |
| -N | 25 | 9.2 | 7.5 | 5.9 | 4.2 | 3.7 |
| -N | 50 | 9.2 | 7.5 | 6.3 | 4.1 | 2.8 |
| +N | 0 | 9.8 | 8.2 | 6.4 | 4.8 | 3.5 |
| +N | 25 | 10.5* | 7.9 | 6.9 | 5.6 | 4.6 |
| +N | 50 | 10.4* | 8.1 | 6.8 | 5.4 | 4.4 |
| +N, nutrients | 0 | 10.2* | 8.3 | 6.5 | 5.0 | 3.6 |
| † Values in parentheses are SOC concentrations prior to study initiation in 2008. * Indicates SOC values for different treatments within a soil depth increment are significantly different at 5% level from initial SOC values in 2008. | ||||||
DownLoad: CSVAs noted previously with SOC concentrations (Table 2), N fertilization and residue return also affected SOC storage differently for continuous and rotated sorghum (Figure 2). Soil organic C content to 90 cm depth increased significantly with time for all treatments in continuous sorghum compared to initial content in 2008. Soil organic C content for rotated sorghum also tended to increase compared to initial values, but differences generally were not significant. Increases in SOC were much higher with continuous than rotated sorghum. For example, SOC storage for unfertilized rotated sorghum averaged across residue return rates increased by 10% (59.4 to 65.1 Mg ha−1) by 2015 and by 9% (59.4 to 64.4 Mg ha−1) for the seven-year average (2009-2015) compared to initial values in 2008. The SOC stock was further increased by about 3% for both 2015 (65.1 to 66.7 Mg ha−1) and seven-year average (64.4 to 66.1 Mg ha−1) values by N fertilization. In contrast, SOC storage to 90 cm for continuous sorghum with N fertilization averaged across residue rates increased by 51% (59.4 to 90.1 Mg ha−1) by 2015 and by 50% (59.4 to 88.5 Mg ha−1) for the seven-year average (2009-2015) compared to initial values. The largest observed SOC content by 2015 was approximately 95 Mg ha−1 and was associated with continuous sorghum, 25% residue return, and N fertilization, and represented a 60% increase (59.4 to 95.2 Mg ha−1) from initial levels. This amount was approximately 12 and 5 Mg ha−1 higher than SOC contents with either 0 or 50% residue return, respectively. In comparison, SOC storage to 90 cm for rotated sorghum also receiving N and 25% residue return was about 30 Mg ha−1 lower than for continuous sorghum after seven years. Significantly increased SOC storage was noted in 2009 after only one year of bioenergy sorghum (data not shown). Meki et al. [12] using a simulation model estimated that under conventional grain sorghum production for 25 years in south central Texas, as much as 130 Mg SOC ha−1 could be stored. Meki et al. [13] also modeled bioenergy sorghum systems and projected that with conventional tillage after 51 years with 25 and 50% residue return, SOC would increase by 16% and 59% compared with 0% return. These authors further estimated that 75% of bioenergy sorghum biomass could potentially be removed from a continuous no till system without any detrimental effect on SOC storage. Dou et al. [56] using the DNDC model, suggested that SOC sequestration will increase with time under bioenergy sorghum receiving residue return and the management practices of the current study.
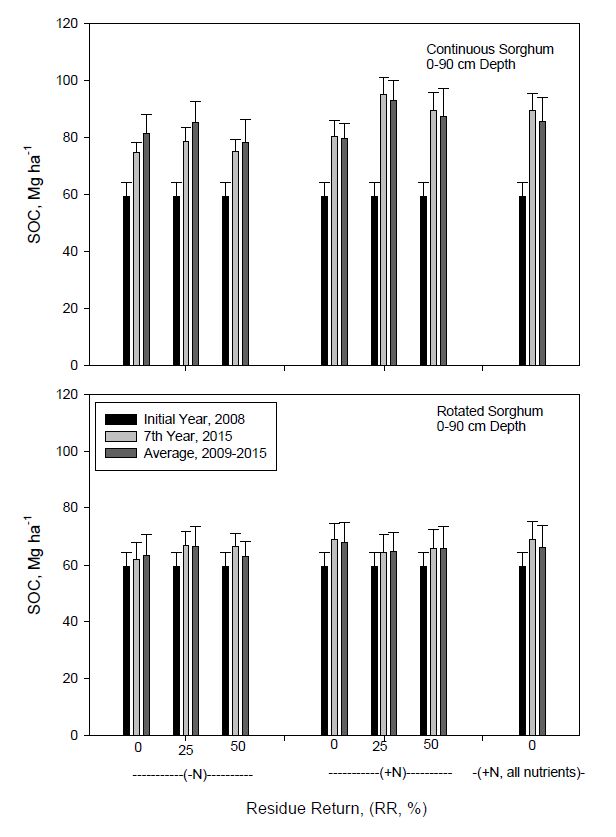 Figure 2. Soil organic carbon (SOC) content to 90 cm depth in continuous and rotated sorghum after seven years compared to initial and the seven year average as influenced by N fertilization and biomass return rate. All nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.
Figure 2. Soil organic carbon (SOC) content to 90 cm depth in continuous and rotated sorghum after seven years compared to initial and the seven year average as influenced by N fertilization and biomass return rate. All nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.Total soil N to 90 cm depth tended to follow similar patterns as those observed for SOC storage (Figure 3). For example, the largest increase in TSN occurred with 25% residue return and N fertilization in continuous sorghum, similar to that for SOC. Total soil N averaged across residue return rate after 7 years of continuous sorghum increased by 29% (9.2 to 11.9 Mg ha−1) and 18% (9.2 to 10.9 Mg ha−1) with or without N fertilization, respectively. The increase in TSN for rotated sorghum was 23% (9.2 to 11.3 Mg ha−1) and 21% (9.2 to 11.1 Mg ha−1) with and without N fertilization. Although quantities of sorghum biomass C returned annually were higher for continuous vs. rotated sorghum (Table 1), amounts of biomass N were similar because of greater N concentrations in rotated sorghum biomass, and may account for similar TSN sequestration over time. Zhang et al. [63] reported that SOC and TSN storage to 60-cm depth for continuous corn with varying straw addition ranged from 45 to 50 and 4.5 to 4.8 Mg ha−1, respectively.
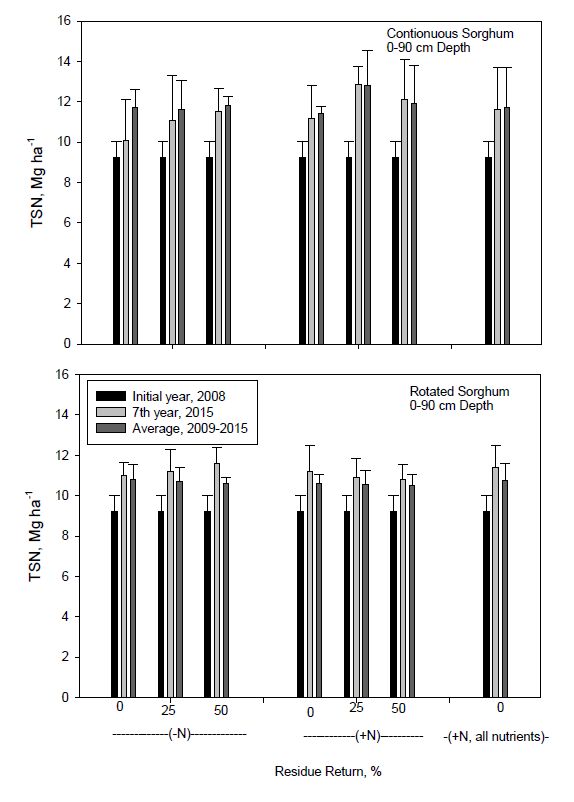 Figure 3. Total soil nitrogen (TSN) content to 90 cm depth in continuous and rotated sorghum after seven years compared to initial and the seven year average as influenced by N fertilization and biomass return rate. All nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.
Figure 3. Total soil nitrogen (TSN) content to 90 cm depth in continuous and rotated sorghum after seven years compared to initial and the seven year average as influenced by N fertilization and biomass return rate. All nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.Soil organic C sequestration was also analyzed by depth to determine where major changes in storage occurred (Figures 4 and 5). For both continuous and rotated sorghum, changes in SOC stocks were greater in lower depths, principally at 30-60 and 60-90 cm and especially for continuous sorghum. Carbon sequestration increased with depth partly, but not totally, due to increased thickness of the sampled soil layer. Continuous sorghum showed a trend for increasing SOC with more residue return for both N regimes at depths to 15 cm (Figure 4). No residue return, however, resulted in slightly greater SOC storage at depths greater than 15 cm compared to residue added treatments in continuous sorghum without N fertilization.
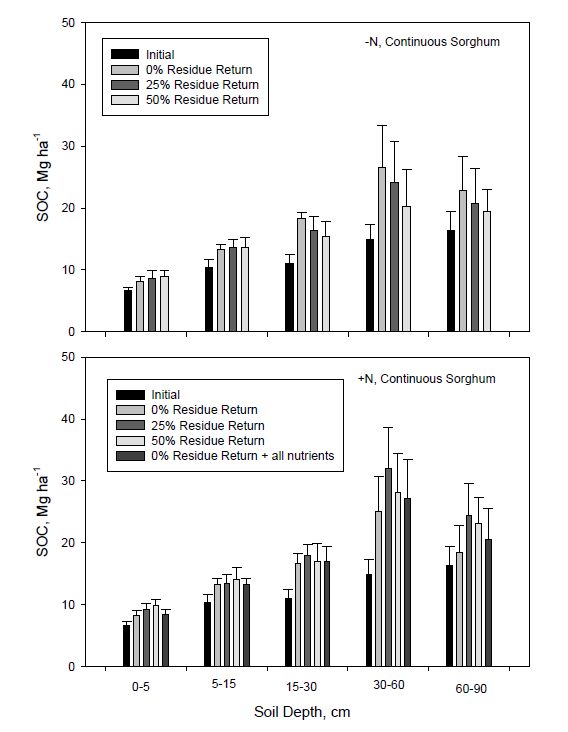 Figure 4. Average soil organic C (SOC) storage to 90-cm depth compared with initial SOC storage in 2008 for continuous sorghum as influenced by N fertilization and residue return rate. All nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.
Figure 4. Average soil organic C (SOC) storage to 90-cm depth compared with initial SOC storage in 2008 for continuous sorghum as influenced by N fertilization and residue return rate. All nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.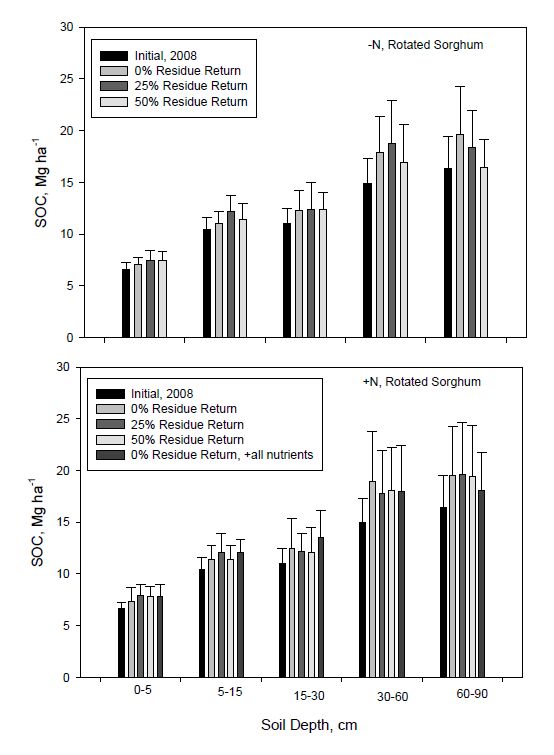 Figure 5. Average soil organic C (SOC) storage to 90-cm depth compared with initial SOC storage in 2008 for rotated sorghum as influenced by N fertilization and residue return rate. All nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.
Figure 5. Average soil organic C (SOC) storage to 90-cm depth compared with initial SOC storage in 2008 for rotated sorghum as influenced by N fertilization and residue return rate. All nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.In contrast, with N fertilization, 25% residue return resulted in greater SOC sequestration compared with other return rates below 15 cm, but especially between 30 and 60 cm. Treatment trends for rotated sorghum (Figure 5) were not as consistent nor differences as large as for continuous sorghum. Increases in SOC for rotated sorghum averaged across residue return rates at 30 to 90 cm depth compared to initial SOC content were about 2 and 4 Mg ha−1 without and with added N, respectively, while changes for continuous sorghum were approximately 2 and 15 Mg ha−1 for these same two N treatments (Figures 4 and 5). With 25% residue return, SOC storage at 30-60 cm depth for continuous sorghum increased by 107% (14.9 to 30.8 Mg ha−1) and 67% (14.9 to 24.9 Mg ha−1) with and without N fertilization, respectively. At 60-90 cm depth, SOC increased by 48% (16.3 to 24.1 Mg ha−1) and 25% (16.3 to 20.4 Mg ha−1) with these same treatments. Soil organic C contents at 30-60 cm depth for rotated sorghum with 25% residue return increased by 19% (14.9 to 17.8 Mg ha−1) and 26% (14.9 to 18.8 Mg ha−1) with and without fertilization, respectively. At 60-90 cm for rotated sorghum with 25% residue return, SOC increased by 19% (16.3 to 19.4 Mg ha−1) and 12% (16.3 to 18.3 Mg ha−1) with and without N fertilization, respectively. The SOC stock at all depths was lower in rotated than continuous sorghum for all treatments.
In general, the results for SOC and TSN (Figures 2 to 5) differ somewhat from findings of residue return for other crops, partly because changes in these characteristics usually take time and should not be evaluated only in surface soil but at deeper depths [21,64,65]. With 50% residue return in our study, the total mean cumulative input of harvested biomass after 7 years for continuous sorghum was ~60 Mg ha−1 with and ~40 Mg ha−1 without N, while for rotated sorghum, returned sorghum biomass was ~34 and 28 Mg ha−1 with and without N fertilization (Table 1). Even though the residue contribution with N fertilization for the two cropping sequences was considerably different, the increase in SOC stock at 0-15 cm depth was relatively the same [7% (22.5 to 24.1 Mg ha−1) and 7% (18.8 to 20.2 Mg ha−1) for continuous and rotated sorghum, respectively (Figures 4 and 5)]. However, with 50% residue return, N addition increased SOC storage for continuous sorghum at 30-60 and 60-90 cm depths by 39% (20.2 to 28.1 Mg ha−1) and 24% (19.4 to 24.1 Mg ha−1), respectively, compared to no N addition, while for rotated sorghum, SOC increased by only 10% (16.4 to 18.1 Mg ha−1) and 15% (16.8 to 19.4 Mg ha−1) at these depths. Even 0% residue return, especially with continuous sorghum, tended to increase SOC storage at all soil depths, but particularly between 15 and 60 cm, compared to initial values and may imply that most SOC increases at these depths were associated with bioenergy sorghum roots. Dalzell et al. [28] suggested that SOC depends on crop root input and rooting depth.Follett et al. [19] found for both switchgrass and corn that over 50% of the increase in SOC was below 30 cm. Muller et al. [66] quantified corn derived-C transfer to soil by stable isotope analysis and concluded that corn root C input was three-fold higher than total shoot C input, while Menichetti et al. [67] also suggested that corn root-derived SOC contributed more to SOC than above-ground crop residues. Ontl et al. [68] showed that root inputs associated with annual bioenergy sorghum and corn feed stocks influenced SOC, while Bai et al. [69] indicated that roots are a major source of organic C input into soil and that root dynamics play a primary role in regulating C- and nutrient-cycling in soil. Liebig et al. [43] reported that bioenergy sorghum, like switchgrass and alfalfa, has a deep root system, and Richter et al. [70] suggested that deep, extensive root systems of bioenergy crops might result in greater soil C sequestration. Meki et al. [13] used simulation modeling to estimate that bioenergy sorghum roots contributed 24, 37, 54, and 95% of total crop residue input, respectively, when above-ground biomass was removed at rates of 0%, 50%, 75%, and 100%. In other words, root biomass becomes more important to SOC when no residue is returned to soil. Bioenergy sorghum may potentially produce 7.0 to 10.7 Mg ha−1 of root biomass [13]. In our study, increased SOC of surface soil may mostly have come from returned residue, while that in the subsurface likely was primarily associated with plant roots.
Water deficit can also alter relative shoot and root productivity. At least two (2011 and 2013) of the seven growing seasons in our study experienced significant rainfall deficit, with 2011 being the most severe drought year on record for the area [24]. Root/shoot ratio tends to increase with drought, largely due to a stronger effect of water deficit on shoot than root growth. In some cases, plant root systems may even continue to expand at very low water potentials, whereas above ground growth is completely halted [71].
Effects of different residue return rates on changes in SOC and TSN stocks with depth after seven years of continuous or three years of rotated bioenergy sorghum receiving N fertilization and are shown in Figures 6 and 7. With N fertilization, the addition of harvested sorghum biomass increased SOC within all depth increments for both sorghum cropping sequences (Figure 6). The nutrient return treatment also increased SOC at all depths compared to complete biomass removal for continuous sorghum and to 30 cm for rotated sorghum. The largest gains in SOC storage occurred with 25% residue return at soil depths of 0-60 and 0-90 cm for continuous sorghum cropping. After seven years of this treatment, SOC increased by 1.75, 5.25, 15.4, 32.9, and 39.2 Mg ha−1 at 0-5, 0-15, 0-30, 0-60, and 0-90 cm depths, respectively. Based on the results, the estimated annual SOC gains with 25% residue return for continuous sorghum were 0.25, 0.75, 2.2, 4.7 and 5.6 Mg ha−1yr−1 at 0-5, 0-15, 0-30, 0-60, and 0-90 cm depths, respectively. The average annual quantity of biomass C returned for N fertilized continuous sorghum at 25% residue return was 2.4 Mg ha−1 yr−1 (Table 1). If one assumes, that at most, 50% of the returned biomass C was sequestered in soil, this amount (1.2 Mg ha−1 C yr−1) only accounts for C sequestered at the 0-15 cm depth and part of the C sequestration at 0-30 cm (Figure 6). The remaining SOC at this depth and below, therefore, was likely associated with roots.
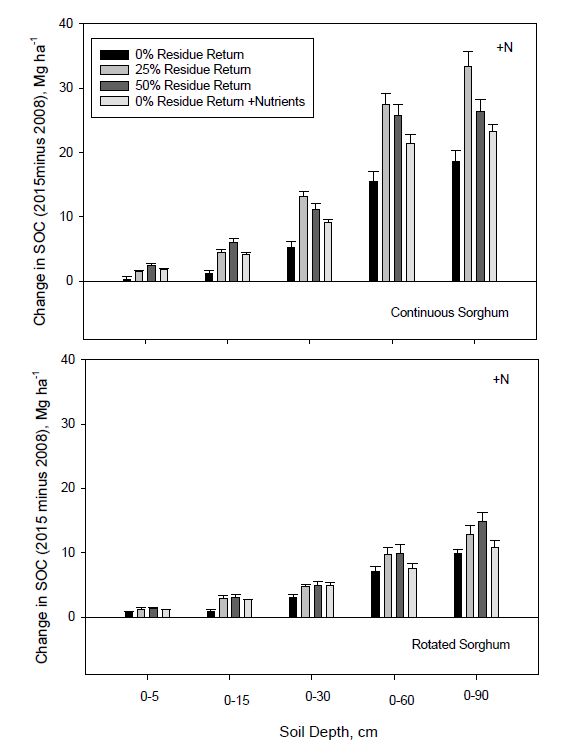 Figure 6. Change in soil organic C (SOC) after 7 years of continuous or 3 years of rotated bioenergy sorghum receiving N fertilization as influenced by residue return rate. +Nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.
Figure 6. Change in soil organic C (SOC) after 7 years of continuous or 3 years of rotated bioenergy sorghum receiving N fertilization as influenced by residue return rate. +Nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.Treatments with rotated sorghum also showed no loss in SOC after seven years and the largest gain occurred with 50% residue return to 90 cm depth (Figure 6). With 50% residue return, the SOC gain for 0-5 cm was 1.4 Mg ha−1 and increased to 3.0, 4.9, 9.9, and 14.8 Mg ha−1 when calculated to 15, 30, 60, and 90 cm, respectively. Estimated annual rates of SOC gain for these same depths were 0.2, 0.4, 0.8, 1.4, and 2.1 Mg ha−1 yr−1. The SOC gain in the surface 0-5 cm was similar for this treatment in both continuous and rotated sorghum, but SOC gain at deeper depths was two to three times higher for continuous compared to rotated sorghum.
Soil organic C sequestration in our experiment was similar to or higher than that reported for several bioenergy crops. Sainju et al. [38] reported that the SOC content for 0-30 cm depth after three years of bioenergy sorghum cropping was 30.5 Mg C ha−1. After seven years of continuous sorghum in our experiment, treatments receiving 0, 25, or 50% residue return and the nutrient return treatment contained 33.2, 39.3, 38.4, and 37.2 Mg C ha−1, respectively, to this same depth. Lemus and Lal [72] reported that bioenergy crops can sequester C at rates ranging from 0 to 3 Mg ha−1 yr−1 in the 0-5 cm depth. Liebig et al. [43] showed that SOC storage by bioenergy crops varied from −0.6 to 4.3 Mg C ha−1 yr−1 and varied both spatially and temporally. Meki’s et al. [13] simulation results indicated that differences in SOC storage with bioenergy sorghum were most apparent in the upper 50 cm of the soil profile and were directly proportional to biomass inputs. In their study, averaged across tillage and cropping treatments, SOC storage gains of 7 and 49% were observed with 50 and 100% biomass return, whereas SOC losses of 15 and 28% were estimated with 25 and 0% return. Our experiment, however, showed no loss of SOC after seven years of bioenergy sorghum cropping with 25% biomass return (Figure 6).
The nutrient return treatment (0% residue return +N + nutrients) also had a positive effect on SOC stock and increased SOC at all measured depths with continuous sorghum, although quantities were lower than those for residue return (Figure 6). Soil organic C increased by 35, 30, 23, 36, and 23% after seven years with this treatment when summed at 5, 15, 30, 60, and 90 cm depths, respectively. Sorghum is known to have a deep rooting pattern that may allow it to capitalize on both nutrient and water uptake with depth [1]. The nutrient return treatment was previously reported to significantly increase P and K availability for bioenergy sorghum production [24]. Blanco-Canqui and Lal [36] reported that greater than 75% stover removal reduced available P and exchangeable K+ by 40 and 15%, respectively.
Total soil N increased at all depths after seven years of treatment regardless of cropping sequence or residue return (Figure 7). Changes in TSN in continuous sorghum generally increased with increasing biomass or nutrient return to 30 cm, but were less consistent at deeper depths, with no return becoming more similar to other treatments. Increases in total N in rotated sorghum were greater than that for continuous sorghum at depths greater than 30 cm, with the 0% return treatment exhibiting similar or greater TSN increases compared to the other treatments at all depths.
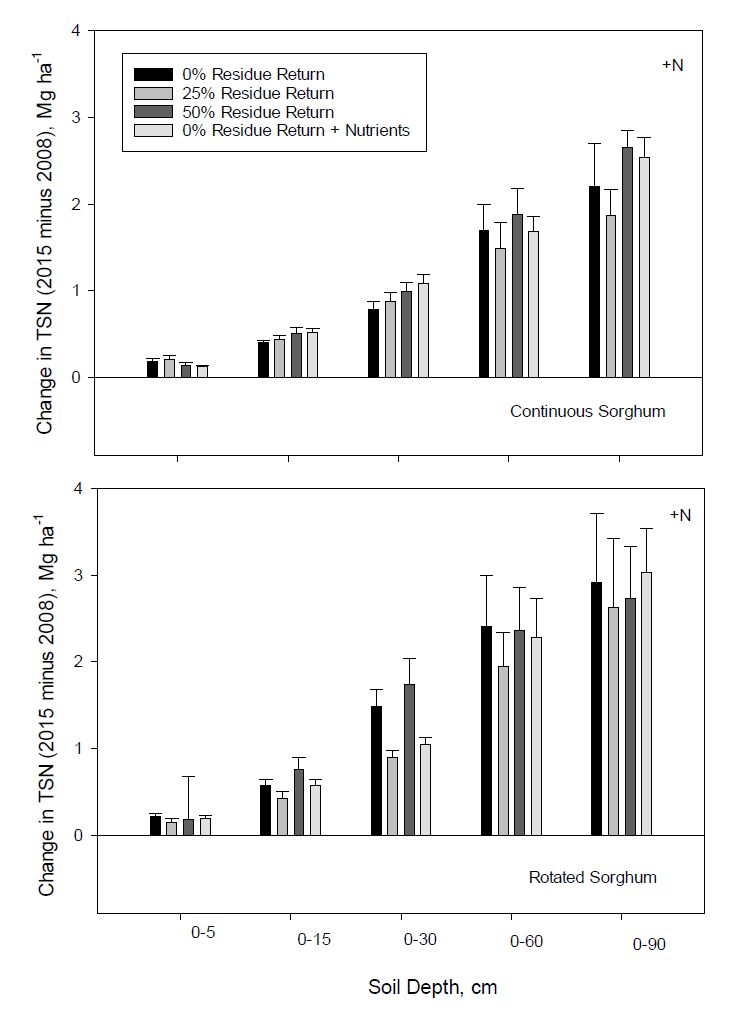 Figure 7. Change in total soil N (TSN) after 7 years of continuous or 3 years of rotated bioenergy sorghum receiving N fertilization as influenced by residue return rate. +Nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.
Figure 7. Change in total soil N (TSN) after 7 years of continuous or 3 years of rotated bioenergy sorghum receiving N fertilization as influenced by residue return rate. +Nutrients represents the nutrient return treatment. Error bars indicate one standard deviation.As would be expected, N in returned biomass was much smaller than associated amounts of organic C. Even if all N returned in biomass over the duration of the study (Table 1) had been conserved in soil, it still couldn’t account for the observed TSN increase even to 15 cm. Thus, N fertilization effects on sorghum growth and root production were likely important contributors to TSN increases at deeper depths. Observed changes in TSN (Figure 7) followed general trends exhibited for SOC (Figure 6), but changes in TSN for individual treatments were more variable. Lal [25] and Khan et al. [73] showed that SOC sequestration rate, especially at lower depths, depended on N availability. Li et al. [74] suggested that increased N might stimulate growth and C accumulation of roots. Hendricks et al. [71], however, found that fine root biomass decreased with increasing N availability. Halvorson et al. [35] and Menichetti et al. [67] reported that N fertilization promoted SOC sequestration. Blanco-Canqui and Lal [36] reported that without N fertilization complete corn stover removal for 4 years reduced total N by 0.82 Mg ha−1 in the 0 to 10 cm depth. Veenstra and Burras [75] found that continuous cropping decreased SOC in surface soil, but increased it with depth.
Returned biomass was assumed to be the main source of SOC and recycled nutrients in this study. However, the extent of aboveground residue or belowground root decomposition relative to the SOC pool potentially depends on their C:N ratios [60], and the C:N ratio of soil organic matter is also somewhat indicative of its stage of decomposition. Li et al. [74] suggested that crop residue with low N concentration and high C:N ratio ( such as corn or sorghum) can be incorporated into soil to immobilize N and potentially decrease CO2 emissions. Soil C:N ratios in our study generally decreased with depth. Ostrowska and Porebska [76] reported a similar pattern, and Langdale et al. [77] and Gal et al. [78] showed that SOC concentration and soil C:N decreased with depth, but were influenced by tillage systems and residue return rate.
Soil C:N under continuous sorghum tended to increase in all depth increments with residue return, but especially in the surface 0-5 cm with N fertilization (Table 3). Nitrogen fertilization and residue return with continuous sorghum also significantly increased soil C:N at 30-60 and 60-90 cm depths compared with initial values. Soil C:N under rotated sorghum also decreased with depth and were lower than those for continuous sorghum, but were not different than initial values. Cai et al [60] concluded that N fertilization affected not only the composition of corn roots, but also their decomposition rate.
| Fertilization | Residue Return | Soil Depth (cm) | ||||
| 0–5 | 5–15 | 15–30 | 30–60 | 60–90 | ||
| % | Continuous Sorghum, C:N ratio | |||||
| -N | 0 | 8.4 (9.1†) | 7.6 (7.9) | 7.5 (7.3) | 6.6 (6.6) | 6.8(6.3) |
| -N | 25 | 9.7 | 8.0 | 8.1 | 7.3 | 6.3 |
| -N | 50 | 12.1* | 8.1 | 7.2 | 6.4 | 6.3 |
| +N | 0 | 9.2 | 7.8 | 7.9 | 7.4 | 6.9 |
| +N | 25 | 10.5* | 8.5 | 7.6 | 7.9* | 8.0* |
| +N | 50 | 10.2* | 8.0 | 7.7 | 7.4 | 7.9* |
| +N, nutrients | 0 | 10.2* | 8.0 | 7.5 | 7.3 | 6.9 |
| Rotated Sorghum, C:N ratio | ||||||
| -N | 0 | 8.9 (9.1†) | 7.5 (7.9) | 6.9 (7.3) | 5.7 (6.6) | 5.5(6.3) |
| -N | 25 | 9.0 | 7.2 | 6.5 | 5.8 | 6.1 |
| -N | 50 | 8.9 | 7.1 | 6.6 | 5.7 | 5.7 |
| +N | 0 | 9.1 | 6.9 | 6.7 | 5.8 | 5.9 |
| +N | 25 | 8.9 | 7.3 | 6.9 | 5.9 | 5.8 |
| +N | 50 | 8.4 | 7.1 | 6.5 | 5.8 | 6.3 |
| +N, nutrients | 0 | 8.7 | 7.2 | 6.4 | 6.2 | 6.2 |
| † Values in parentheses are soil C:N ratios prior to study initiation in 2008. * Indicates soil C:N ratios are significantly different at 5% level from initial C:N ratio values in 2008. | ||||||
DownLoad: CSVThe highest C:N ratios were observed in surface soil under continuous sorghum at 25 and 50% residue return rates with or without N fertilization (Table 3) and may have resulted from returning relatively large quantities of biomass with relatively high C:N ratios. Increased soil C:N at deeper depths in continuous sorghum was probably associated with sorghum roots. Corn and sorghum root C:N ratios have been reported to be around 50:1 and 80:1, respectively [56]. Wilhelm et al. [7] stated that the greater contribution of roots to SOC than shoots was attributed to roots having greater lignin and lower soluble C compared with shoots. Initial decomposition of crop residue correlated with soluble C, but later decomposition correlated with the concentration of cellulose, hemicelluloses, and lignin.
Based on SOC and TSN from various biomass sorghum residue return rates with N fertilization, 75% removal of bioenergy sorghum biomass would appear to be feasible for maintaining soil quality of the soil under investigation in the long term. Harvesting 75% and returning 25% of residue without detrimentally affecting SOC storage agreed with previous crop model results [13,56]. Nitrogen fertilization was essential for maintaining high biomass yield over time, especially with continuous sorghum. For bioenergy sorghum grown in rotation with corn, biomass yield was maintained over time with N fertilization, but minimal sorghum residue return. Soil organic C and TSN increased with time for both continuous and rotated biomass sorghum, but sequestration was higher for continuous. For continuous sorghum after seven years of N fertilization and 25% biomass return, SOC increased by 1.75, 5.25, 15.4, 32.9, and 39.2 Mg ha−1, respectively, when summed over 0-5, 0-15, 0-30, 0-60, and 0-90 cm depths. The largest SOC gains for rotated sorghum occurred with N fertilization and 50% residue return, amounting to 1.4, 3.0, 4.9, 9.9, and 14.8 Mg ha−1 when calculated to 5, 15, 30, 60, and 90 cm depths, respectively. Relatively greater sequestration with depth may indicate the importance of biomass sorghum roots for SOC sequestration.
This research was supported by the Texas A&M AgriLife Research Cropping Systems Program and the USDA/NIFA Agriculture and Food Research Initiative (grant #2022-67009-30050).
All authorsdeclare no conflict of interests in this paper.
| [1] | Rooney WL, Blumenthal J, Bean B, et al. (2007) Designing sorghum as a dedicated bioenergy feedstock. Biofuel Bioprod Bior 1: 147−157. |
| [2] |
Manatt RK, Hallam A, Schulte LA, et al. (2013) Farm-scale costs and returns for second generation bioenergy cropping systems inthe US Corn Belt. Environ Res Letters 8: 035037. doi: 10.1088/1748-9326/8/3/035037

|
| [3] | Storlien JO, Hons FM, Wight JP, et al. (2014) Carbon dioxide and nitrous oxide emissions impacted by bioenergy sorghum management. Soil Sci Soc Am J doi: 10.2136/sssaj2014.04.0176. |
| [4] |
Tilman D, Socolow R, Foley JA, et al. (2009) Beneficial biofuels—the food, energy, and environment trilemma. Science 325: 270–271. doi: 10.1126/science.1177970
|
| [5] |
Gill JR, Burks PS, Staggenborg SA et al. (2014) Yield results and stability analysis from the sorghum regional biomass feedstock trial. Bioenerg Res 7: 1026–1034. doi: 10.1007/s12155-014-9445-5
|
| [6] |
Blanco-Canqui H (2013) Crop residue removal for bioenergy reduces soil carbon pools: How can we offset carbon losses. Bioenerg Res 6: 358–371. doi: 10.1007/s12155-012-9221-3
|
| [7] |
Wilhelm WW, Johnson JM, Hatfield JL, et al. (2004) Crop and soil productivity response to corn residue removal: a literature review. Agron J 96: 1–17. doi: 10.2134/agronj2004.0001
|
| [8] |
Gollany HT, Rickman RW, Liang Y, et al. (2011) Predicting agricultural management influence on long-term soil organic carbon dynamics: implications for biofuel production. Agron J 103: 234–24618. doi: 10.2134/agronj2010.0203s
|
| [9] | Muth DJ, Bryden KM, Nelson RG (2013) Sustainable agricultural residue removal for bioenergy: A spatially comprehensive U.S. national assessment. Appl Energy 102: 403–417. |
| [10] |
Stewart CE, Follett RF, Pruessner EG, et al. (2015) Nitrogen and harvest effects on soil properties under rainfed switchgrass and no-till corn over 9 years: Implications for soil quality. GCB Bioenergy 7: 288–301. doi: 10.1111/gcbb.12142
|
| [11] | USDA NRCS (USDA Natural Resources Conservation Service) (2006) Crop residue removal for biomass production: effects on soils and recommendations. Soil quality-Agronomy Technical Note No. 19. |
| [12] | Meki MN, Kemanian AR, Potter SR, et al. (2013a) Cropping system effects on sorghum grain yield, soil organic carbon, and global warming potential in central and south Texas. Agr Systems 117: 19–29. |
| [13] | Meki MN, Snider JL, Kiniry JR, et al. (2013b) Energy sorghum biomass harvest threshold and tillage effects on soil organic carbon and bulk density. Ind Crops Prod 43: 172–182. |
| [14] |
Reijnders L (2013) Sustainability of soil fertility and the use of lignocellulosic crop harvest residues for the production of biofuels: a literature review. Environ Tech 34: 1725–1734. doi: 10.1080/09593330.2013.826252
|
| [15] | Nelson RG, Tatarko J, Ascough JC II (2015) Soil erosion and organic matter variations for central Great Plains cropping systems under residue removal. Transactions ASABE 58: 415–427. |
| [16] |
Reeves DW (1997) The role of soil organic matter in maintaining soil quality in continuous cropping systems. Soil Till Res 43: 131–167. doi: 10.1016/S0167-1987(97)00038-X
|
| [17] | Liu X, Herbert SJ, Hashemi AM, et al. (2006) Effects of agricultural management on soil organic matter and carbon transformation—a review. Plant Soil Environ 52: 531–543. |
| [18] |
Baker JM, Ochsner TE, Venterea RT, et al. (2007) Tillage and soil carbon sequestration - What do we really know? Agr Ecosyst Environ 118: 1–5. doi: 10.1016/j.agee.2006.05.014
|
| [19] |
Follett RF, Vogel KP, Varvel GE, et al. (2012) Soil carbon sequestration by switchgrass and no-till maize grown for bioenergy. Bioenerg Res 5: 866–875. doi: 10.1007/s12155-012-9198-y
|
| [20] |
Gollany HT, Novak JM, Liang Y, et al. (2010) Simulating soil organic carbon dynamics with residue removal using the CQESTR model. Soil Sci Soc Am J 74: 372–383. doi: 10.2136/sssaj2009.0086
|
| [21] |
West TO, Post W (2002) Soil organic carbon sequestration rates by tillage and crop rotation: A global data analysis. Soil Sci Soc Am J 66: 1930–1946. doi: 10.2136/sssaj2002.1930
|
| [22] |
Dou F, Hons FM (2006) Tillage and nitrogen effects on soil organic matter fractions in wheat-based systems. Soil Sci Soc Am J 70: 1896–1905. doi: 10.2136/sssaj2005.0229
|
| [23] |
Dou F, Wright AL, Hons FM (2008) Sensitivity of labile soil organic carbon to tillage in wheat-based cropping systems. Soil Sci Soc Am J 72: 1445–1453. doi: 10.2136/sssaj2007.0230
|
| [24] |
Shahandeh H, Hons FM, Wight JP, et al. (2015) Harvest strategy and N fertilizer effects on bioenergy sorghum production. AIMS Energy 3: 377–400. doi: 10.3934/energy.2015.3.377
|
| [25] |
Lal R (2009) Soil quality impacts of residue removal for bioethanol production. Soil Till Res 102: 233–241. doi: 10.1016/j.still.2008.07.003
|
| [26] |
Somerville C, Youngs H, Taylor C, et al. (2010) Feedstocks for lignocellulosic biofuels. Science 329: 790–792. doi: 10.1126/science.1189268
|
| [27] |
Kirkby CA, Richardson AE, Wade LJ, et al. (2014) Nutrient availability limits carbon sequestration in arable soils. Soil Biol Biochem 68: 402–409. doi: 10.1016/j.soilbio.2013.09.032
|
| [28] |
Dalzell BJ, Johnson JMF, Tallaksen J, et al. (2013) Simulated impacts of crop residue removal and tillage on soil organic matter maintenance. Soil Sci Soc Am J 77: 1349–1356 doi: 10.2136/sssaj2012.0221
|
| [29] |
Blanco-Canquil H, Lal R (2007) Soil and crop response to harvesting corn residues for biofuel production. Geoderma 141: 355–362. doi: 10.1016/j.geoderma.2007.06.012
|
| [30] | Graham RL, Nelson R, Sheehan R, et al. (2007) Current and potential U.S. corn stover supplies. Agron J 99: 1–11. |
| [31] |
Anderson-Teixeira KJ, Davis SC, Masters MD (2009) Changes in soil organic carbon under biofuel crops. GCB Bioenerg 1: 75–96. doi: 10.1111/j.1757-1707.2008.01001.x
|
| [32] | Johnson JM, Novak JM, Varvel GE, et al. ( 2014). Crop residue mass needed to maintain soil organic carbon levels: can it be determined? Bioenerg Res 7: 481–490. |
| [33] |
Wilhelm WW, Johnson JM, Karlen DL, et al. (2007) Corn stover to sustain soil organic carbon further constrains biomass supply. Agron J 99: 1665–1667. doi: 10.2134/agronj2007.0150
|
| [34] |
Johnson JD, Allmaras D, Reicosky RR (2006) Estimating source carbon residues, root and rhizodeposits using the national grain—yield database. Agron J 98: 622–636. doi: 10.2134/agronj2005.0179
|
| [35] | HalvorsonAD, WienholdBJ, Black AL (2002) Tillage, nitrogen, and cropping system effects on soil carbon sequestration. Soil Sci Soc Am J 66 : 906–912. |
| [36] | Blanco-Canqui H, Lal R (2009) Crop residue management and soil carbon dynamics, In: Lal R, Follett RF, Soil carbon sequestration and the greenhouse effect: SSSA Special Publication 57. 2nd ed. ASA, and SSSA, Madison, WI, 291–309. |
| [37] |
Halvorson AD, Schlege AJ (2012) Crop rotation effect on soil carbon and nitrogen stocks under limited irrigation. Agron J 104: 1265–1273. doi: 10.2134/agronj2012.0113
|
| [38] | Sainju UM, Singh HP, Singh BP (2015) Cover crop effects on soil carbon and nitrogen under bioenergy sorghum crops. J Soil Water Conserv 70: 410–417. |
| [39] | Anderson-Teixeria KJ, Masters MD, Black CK, et al. (2013) Altered belowground carbon cycling, following land-use change to perennial bioenergy crops. Ecosyst DOI 10. 1007/s10021-021-9628-x. |
| [40] |
Mazzilli SR, Kemanian AR, Ernst OR, et al. (2015) Greater humification of belowground than aboveground biomass carbon into particulate soil organic matter in no-till corn and soybean crops. Soil Biol Biochem 85: 22–30. doi: 10.1016/j.soilbio.2015.02.014
|
| [41] |
Flesa H, Ludwiso B, Heil B, et al. (2000) The origin of soil organic C, dissolved organic C and respiration in a long term maize experiment in Halle, Germany determined by 13C natural abundance. J Plant Nutr Soil Sci 163: 157–163. doi: 10.1002/(SICI)1522-2624(200004)163:2<157::AID-JPLN157>3.0.CO;2-9
|
| [42] |
Wilts AR, Reicosky DC, Allmaras RR, et al. (2004) Long-term corn residue effects: harvest alternatives, soil carbon turnover, and root-derived carbon. Soil Sci Soc Am J 68: 1342–1351. doi: 10.2136/sssaj2004.1342
|
| [43] |
Liebig MA, Schmer MR, Vogel KP, et al. (2008) Soil carbon storage by switchgrass grown for bioenergy. Bioenerg Res 1: 215–222. doi: 10.1007/s12155-008-9019-5
|
| [44] |
Schmidt MWI, Torn MS, Abiven S, et al. (2011) Persistence of soil organic matter as an ecosysytem property. Nature 478: 49–56. doi: 10.1038/nature10386
|
| [45] |
Wight JP, Hons FM, Storlien JO, et al. (2012). Management effects on bioenergy sorghum growth, yield and nutrient uptake. Biomass Bioenerg 46: 593–604. doi: 10.1016/j.biombioe.2012.06.036
|
| [46] |
Adams, WA (1973) The effect of organic matter on the bulk and true densities of some uncultivated podzolic soils. J Soil Sci 24: 10–17. doi: 10.1111/j.1365-2389.1973.tb00737.x
|
| [47] |
Franzluebbers AJ, Hons FM, Zuberer DA (1995) Tillage and crop effects on seasonal dynamics of soil CO2 evolution, water content, temperature, and bulk density. Appl Soil Ecol 2: 95–109. doi: 10.1016/0929-1393(94)00044-8
|
| [48] | Schulte EE, Hopkins BG (1996) Estimation of soil organic matter by weight loss-on-ignition. In Soil organic matter: analysis and interpretation. Madison, WI, U.S.A. Soil Sci Soc Am p 21–27. |
| [49] | Hons FM, Wright AL, Kolodziej SM, et al., (2004) Rotation, tillage, and nitrogen rate effects on cotton growth and yield. In: Proceedings of Annual Beltwide Cotton Conference. January 2004, San Antonio, TX. |
| [50] |
Barber SA (1972) Relation of weather to the influence of hay crops on subsequent corn yields on a Chalmers silt loam. Agron J 64: 8–10. doi: 10.2134/agronj1972.00021962006400010003x
|
| [51] | Cook RJ (1984) Root health: Importance and relationship to farming practices. In Organic Farming: Current Technology and Its Role in a Sustainable Agriculture. Eds. Bezdicek DF, Powers JF. pp 111-127. Am. Soc. Agron. Special Publ. 46. Am. Soc. Agron., Madison, WI. |
| [52] |
Jawson MD, Franzluebbers AJ, Galusha DK, et al. (1993) Soil fumigation within monoculture and rotations: Response of corn and mycorrhizae. Agron J 85: 1174–1180. doi: 10.2134/agronj1993.00021962008500060016x
|
| [53] | Turco R F, Bischoff M, Breakwell DP, et al. (1990) Contribution of soil-borne bacteria to the rotation effect in corn. Plant Soil 122: 115–120. |
| [54] |
Whiting KR, Crookston RK (1993) Host-specific pathogens do not account for the corn-soybean rotation effect. Crop Sci 33: 539–543. doi: 10.2135/cropsci1993.0011183X003300030024x
|
| [55] | Weston LA, Alsaadawi IS, Baerson SR (2013) Sorghum allelopathy—from ecosystem to molecule. J Chem Ecol DOI 10.1007/s10886-013-0245-8. |
| [56] | Dou F, Wight JP, Wilson LT, Storlien JO, et al. (2014) Simulation of biomass yield and soil organic carbon under bioenergy sorghum production. PLoS ONE 9(12): e115598. doi: 10.1371/journal.pone.0115598. |
| [57] | Hao B, Xue Q, Bean BW, et al. (2014) Biomass production, water and nitrogen use efficiency in photoperiod-sensitive sorghum in the Texas High Plains. Biomass Bioenerg 62: 108–116. |
| [58] |
Cadoux S, Ferchaud F, Demay C, et al. (2014) Implication of productivity and nutrient requirements on greenhouse gas balance of annual and perennial bioenergy crops. GCB Bioenerg 6: 425–438. doi: 10.1111/gcbb.12065
|
| [59] |
Halvorson AD, Stewart CE (2015) Stover removal affects no-till irrigated corn yields, soil carbon, and nitrogen. Agron J 107: 1504–1512. doi: 10.2134/agronj15.0074
|
| [60] | Cai M, Dong Y, Chen Z, et al. (2015) Effects of nitrogen fertilizer on the composition of maize roots and their decomposition at different soil depths. Eur J Soil Biol 67: 43–50.69. |
| [61] |
Follett RF (2001) Soil management concepts and carbon sequestration in cropland soils. Soil Till Res 61: 77–92. doi: 10.1016/S0167-1987(01)00180-5
|
| [62] |
Franzluebbers AJ (2010) Achieving soil organic carbon sequestration with conservation agricultural systems in the Southeastern United States. Soil Sci Soc Am J 74: 347–357. doi: 10.2136/sssaj2009.0079
|
| [63] |
Zhang P, Wei T, Li Y, et al. (2015) Effects of straw incorporation on stratification of the soil organic C, total N and C: N ratio in a semiarid region of China. Soil Till Res 153: 28–35. doi: 10.1016/j.still.2015.04.008
|
| [64] |
Varvel GE, Wilhelm WW (2011) No-tillage increases soil profile carbon and nitrogen under long-term rainfed cropping systems. Soil Till Res 114: 28–36. doi: 10.1016/j.still.2011.03.005
|
| [65] |
Clapp, CE, Allmaras RR, Layese ME, et al. (2000) Soil organic carbon and 13C abundance as related to tillage, crop residue, and nitrogen fertilization under continuous corn management in Minnesota. Soil Till Res 55: 127–142. doi: 10.1016/S0167-1987(00)00110-0
|
| [66] |
Muller K, Kramer S, Haslwimmer H (2016) Carbon transfer from maize roots and litter into bacteria and fungi depends on soil depth and time. Soil Biol Biochem 93: 79–89. doi: 10.1016/j.soilbio.2015.10.015
|
| [67] |
Menichetti L, Ekblad A, Katterer T (2015) Contribution of roots and amendments to soil carbon accumulation within the soil profile in a long-term field experiment in Sweden. Agr Ecosyst and Environ 200: 79–87. doi: 10.1016/j.agee.2014.11.003
|
| [68] |
Ontl AT, Hofmockel KS, Cambardella, CA, et al. (2013) Topographic and soil influences on root productivity of three bioenergy cropping systems. New Phytologist 199: 727–737. doi: 10.1111/nph.12302
|
| [69] |
Bai WM, Zhou M, Fang Y, et al. (2015) Differences in spatial and temporal root lifespan of temperate steppes across Inner Mongolia grasslands. Biogeosciences Discuss 12: 19999–20023. doi: 10.5194/bgd-12-19999-2015
|
| [70] |
Richter GM, Agostini F, Redmile-Gordon M, et al. (2015) Sequestration of C in soils under Miscanthus can be marginal and is affected by genotype-specific root distribution. Agr Ecosyst Environ 200: 169–177. doi: 10.1016/j.agee.2014.11.011
|
| [71] |
Hendricks JJ, Hendrick RL, Wilson CA, et al. (2006) Assessing the patterns and controls of fine root dynamics: an empirical test and methodological review. J Ecol 94: 40–57. doi: 10.1111/j.1365-2745.2005.01067.x
|
| [72] | Lemus R, Lal R (2005): Bioenergy crops and carbon sequestration. Crit Rev Plant Sci 24: 1–21. |
| [73] |
Khan SA, Mulvaney RL, Ellsworth TR, et al. (2007) The myth of nitrogen fertilization for soil carbon sequestration. J Environ Qual 36: 1821–1832. doi: 10.2134/jeq2007.0099
|
| [74] |
Li W, Jin C, Guan D, et al. (2015) The effects of simulated nitrogen deposition on plant root traits: a meta-analysis. Soil Biol Biochem 82: 112–118. doi: 10.1016/j.soilbio.2015.01.001
|
| [75] |
Veenstra JJ, Burras CL (2015) Soil profile transformation after 50 years of agricultural land use. Soil Sci Soc Am J 79: 1154–1162. doi: 10.2136/sssaj2015.01.0027
|
| [76] |
Ostrowska A, Porebska G (2015) Assessment of the C/N ratio as an indicator of the decomposability of organic matter in forest soils. Ecol Indicators 49: 104–109. doi: 10.1016/j.ecolind.2014.09.044
|
| [77] | Langdale GW, Hargrove WL, Giddens J (1984) Residue management in double-crop conservation tillage systems. Agron J 76: 689–694. |
| [78] |
Gal A, Vyn TJ, Micheli E, et al. (2007) Soil carbon and nitrogen accumulation with long term no till versus moldboard plowing overestimated with tilled-zone sampling depths. Soil Till Res 96: 42–51. doi: 10.1016/j.still.2007.02.007
|
| 1. | Brian A. McKinley, Sara N. Olson, Kimberley B. Ritter, Dustin W. Herb, Steven D. Karlen, Fachuang Lu, John Ralph, William L. Rooney, John E. Mullet, Prasanta K. Subudhi, Variation in energy sorghum hybrid TX08001 biomass composition and lignin chemistry during development under irrigated and non-irrigated field conditions, 2018, 13, 1932-6203, e0195863, 10.1371/journal.pone.0195863 | |
| 2. | Ka Man Jasmine Yu, Joel Oliver, Brian McKinley, Brock Weers, Hilary T. Fabich, Nathan Evetts, Mark S. Conradi, Stephen A. Altobelli, Amy Marshall‐Colon, John Mullet, Bioenergy sorghum stem growth regulation: intercalary meristem localization, development, and gene regulatory network analysis, 2022, 112, 0960-7412, 476, 10.1111/tpj.15960 | |
| 3. | Austin Lamb, Brock Weers, Brian McKinley, William Rooney, Cristine Morgan, Amy Marshall‐Colon, John Mullet, Bioenergy sorghum’s deep roots: A key to sustainable biomass production on annual cropland, 2022, 14, 1757-1693, 132, 10.1111/gcbb.12907 | |
| 4. | Austin Lamb, Evan Kurtz, Priscilla Glenn, Brian A. McKinley, John Mullet, Bioenergy sorghum nodal root bud development: morphometric, transcriptomic and gene regulatory network analysis, 2024, 15, 1664-462X, 10.3389/fpls.2024.1456627 | |
| 5. | Emily R. Murray, Jeremiah J. Minich, Jocelyn Saxton, Marie de Gracia, Nathaniel Eck, Nicholas Allsing, Justine Kitony, Kavi Patel-Jhawar, Eric E. Allen, Todd P. Michael, Nadia Shakoor, Philips O. Akinwole, Azdayanti Muslim, Soil depth determines the microbial communities in Sorghum bicolor fields within a uniform regional environment , 2025, 2165-0497, 10.1128/spectrum.02928-24 | |
| 6. | Ka Man Jasmine Yu, Brock D. Weers, Brian A. McKinley, Priscilla D. Glenn, Evan Kurtz, William L. Rooney, John E. Mullet, Bioenergy sorghum stem density increases threefold following internode elongation due to continued accumulation of lignified cell walls and complex regulation of genes involved in cell wall biosynthesis, 2025, 18, 2731-3654, 10.1186/s13068-025-02659-w |







Figures(7) / Tables(3)

Hamid Shahandeh, Frank M. Hons, Joseph O. Storlien, Jason P. Wight. Long-term bioenergy sorghum harvest strategy and soil quality[J]. AIMS Energy, 2016, 4(4): 633-657. doi: 10.3934/energy.2016.4.633

| Fertilization | Residue Return | Continuous Sorghum | Rotated Sorghum | ||||||
| Biomass | C | N | C:N | Biomass | C | N | C:N | ||
| % | ---Mg ha-1 | kg ha-1 | ---Mg ha-1 | kg ha-1 | |||||
| Total Return | |||||||||
| -N | 25 | 21.0 | 8.7 | 162 | 54 | 14.6 | 6.1 | 145 | 42 |
| -N | 50 | 39.7 | 16.4 | 288 | 57 | 28.2 | 11.8 | 284 | 42 |
| +N | 25 | 34.5 | 14.4 | 318 | 45 | 16.7 | 6.9 | 217 | 32 |
| +N | 50 | 59.9 | 24.8 | 530 | 47 | 33.6 | 14.1 | 423 | 33 |
| Annual Return † | |||||||||
| -N | 25 | 3.5 | 1.5 | 27 | 55 | 2.4 | 1.0 | 24 | 42 |
| -N | 50 | 6.6 | 2.7 | 48 | 56 | 4.7 | 2.0 | 47 | 41 |
| +N | 25 | 5.8 | 2.4 | 53 | 45 | 2.8 | 1.2 | 36 | 32 |
| +N | 50 | 10.0 | 4.1 | 88 | 47 | 5.6 | 2.4 | 71 | 33 |
| † Average of 6 years. | |||||||||
DownLoad: CSV| Fertilization | Residue Return | Soil Depth (cm) | ||||
| 0–5 | 5–15 | 15–30 | 30–60 | 60–90 | ||
| % | Continuous Sorghum, SOC, g kg-1 | |||||
| -N | 0 | 9.8 (9.3†) | 8.3 (8.0) | 7.1 (6.5) | 4.5 (4.9) | 3.6(3.7) |
| -N | 25 | 11.6* | 8.5 | 7.3 | 4.9 | 3.7 |
| -N | 50 | 12.0* | 8.9 | 6.2 | 3.9 | 4.0 |
| +N | 0 | 10.1* | 8.1 | 7.4 | 5.7* | 3.7 |
| +N | 25 | 12.1* | 9.0* | 7.6 | 7.3* | 6.7* |
| +N | 50 | 12.8* | 9.4* | 7.1 | 6.2* | 5.6* |
| +N, nutrients | 0 | 10.2* | 9.1* | 7.6 | 6.4* | 4.6 |
| Rotated Sorghum, SOC, g kg-1 | ||||||
| -N | 0 | 9.6 (9.3†) | 7.5 (8.0) | 5.6* (6.5) | 3.9 (4.9) | 3.2(3.7) |
| -N | 25 | 9.2 | 7.5 | 5.9 | 4.2 | 3.7 |
| -N | 50 | 9.2 | 7.5 | 6.3 | 4.1 | 2.8 |
| +N | 0 | 9.8 | 8.2 | 6.4 | 4.8 | 3.5 |
| +N | 25 | 10.5* | 7.9 | 6.9 | 5.6 | 4.6 |
| +N | 50 | 10.4* | 8.1 | 6.8 | 5.4 | 4.4 |
| +N, nutrients | 0 | 10.2* | 8.3 | 6.5 | 5.0 | 3.6 |
| † Values in parentheses are SOC concentrations prior to study initiation in 2008. * Indicates SOC values for different treatments within a soil depth increment are significantly different at 5% level from initial SOC values in 2008. | ||||||
DownLoad: CSV| Fertilization | Residue Return | Soil Depth (cm) | ||||
| 0–5 | 5–15 | 15–30 | 30–60 | 60–90 | ||
| % | Continuous Sorghum, C:N ratio | |||||
| -N | 0 | 8.4 (9.1†) | 7.6 (7.9) | 7.5 (7.3) | 6.6 (6.6) | 6.8(6.3) |
| -N | 25 | 9.7 | 8.0 | 8.1 | 7.3 | 6.3 |
| -N | 50 | 12.1* | 8.1 | 7.2 | 6.4 | 6.3 |
| +N | 0 | 9.2 | 7.8 | 7.9 | 7.4 | 6.9 |
| +N | 25 | 10.5* | 8.5 | 7.6 | 7.9* | 8.0* |
| +N | 50 | 10.2* | 8.0 | 7.7 | 7.4 | 7.9* |
| +N, nutrients | 0 | 10.2* | 8.0 | 7.5 | 7.3 | 6.9 |
| Rotated Sorghum, C:N ratio | ||||||
| -N | 0 | 8.9 (9.1†) | 7.5 (7.9) | 6.9 (7.3) | 5.7 (6.6) | 5.5(6.3) |
| -N | 25 | 9.0 | 7.2 | 6.5 | 5.8 | 6.1 |
| -N | 50 | 8.9 | 7.1 | 6.6 | 5.7 | 5.7 |
| +N | 0 | 9.1 | 6.9 | 6.7 | 5.8 | 5.9 |
| +N | 25 | 8.9 | 7.3 | 6.9 | 5.9 | 5.8 |
| +N | 50 | 8.4 | 7.1 | 6.5 | 5.8 | 6.3 |
| +N, nutrients | 0 | 8.7 | 7.2 | 6.4 | 6.2 | 6.2 |
| † Values in parentheses are soil C:N ratios prior to study initiation in 2008. * Indicates soil C:N ratios are significantly different at 5% level from initial C:N ratio values in 2008. | ||||||
DownLoad: CSV| Fertilization | Residue Return | Continuous Sorghum | Rotated Sorghum | ||||||
| Biomass | C | N | C:N | Biomass | C | N | C:N | ||
| % | ---Mg ha-1 | kg ha-1 | ---Mg ha-1 | kg ha-1 | |||||
| Total Return | |||||||||
| -N | 25 | 21.0 | 8.7 | 162 | 54 | 14.6 | 6.1 | 145 | 42 |
| -N | 50 | 39.7 | 16.4 | 288 | 57 | 28.2 | 11.8 | 284 | 42 |
| +N | 25 | 34.5 | 14.4 | 318 | 45 | 16.7 | 6.9 | 217 | 32 |
| +N | 50 | 59.9 | 24.8 | 530 | 47 | 33.6 | 14.1 | 423 | 33 |
| Annual Return † | |||||||||
| -N | 25 | 3.5 | 1.5 | 27 | 55 | 2.4 | 1.0 | 24 | 42 |
| -N | 50 | 6.6 | 2.7 | 48 | 56 | 4.7 | 2.0 | 47 | 41 |
| +N | 25 | 5.8 | 2.4 | 53 | 45 | 2.8 | 1.2 | 36 | 32 |
| +N | 50 | 10.0 | 4.1 | 88 | 47 | 5.6 | 2.4 | 71 | 33 |
| † Average of 6 years. | |||||||||
| Fertilization | Residue Return | Soil Depth (cm) | ||||
| 0–5 | 5–15 | 15–30 | 30–60 | 60–90 | ||
| % | Continuous Sorghum, SOC, g kg-1 | |||||
| -N | 0 | 9.8 (9.3†) | 8.3 (8.0) | 7.1 (6.5) | 4.5 (4.9) | 3.6(3.7) |
| -N | 25 | 11.6* | 8.5 | 7.3 | 4.9 | 3.7 |
| -N | 50 | 12.0* | 8.9 | 6.2 | 3.9 | 4.0 |
| +N | 0 | 10.1* | 8.1 | 7.4 | 5.7* | 3.7 |
| +N | 25 | 12.1* | 9.0* | 7.6 | 7.3* | 6.7* |
| +N | 50 | 12.8* | 9.4* | 7.1 | 6.2* | 5.6* |
| +N, nutrients | 0 | 10.2* | 9.1* | 7.6 | 6.4* | 4.6 |
| Rotated Sorghum, SOC, g kg-1 | ||||||
| -N | 0 | 9.6 (9.3†) | 7.5 (8.0) | 5.6* (6.5) | 3.9 (4.9) | 3.2(3.7) |
| -N | 25 | 9.2 | 7.5 | 5.9 | 4.2 | 3.7 |
| -N | 50 | 9.2 | 7.5 | 6.3 | 4.1 | 2.8 |
| +N | 0 | 9.8 | 8.2 | 6.4 | 4.8 | 3.5 |
| +N | 25 | 10.5* | 7.9 | 6.9 | 5.6 | 4.6 |
| +N | 50 | 10.4* | 8.1 | 6.8 | 5.4 | 4.4 |
| +N, nutrients | 0 | 10.2* | 8.3 | 6.5 | 5.0 | 3.6 |
| † Values in parentheses are SOC concentrations prior to study initiation in 2008. * Indicates SOC values for different treatments within a soil depth increment are significantly different at 5% level from initial SOC values in 2008. | ||||||
| Fertilization | Residue Return | Soil Depth (cm) | ||||
| 0–5 | 5–15 | 15–30 | 30–60 | 60–90 | ||
| % | Continuous Sorghum, C:N ratio | |||||
| -N | 0 | 8.4 (9.1†) | 7.6 (7.9) | 7.5 (7.3) | 6.6 (6.6) | 6.8(6.3) |
| -N | 25 | 9.7 | 8.0 | 8.1 | 7.3 | 6.3 |
| -N | 50 | 12.1* | 8.1 | 7.2 | 6.4 | 6.3 |
| +N | 0 | 9.2 | 7.8 | 7.9 | 7.4 | 6.9 |
| +N | 25 | 10.5* | 8.5 | 7.6 | 7.9* | 8.0* |
| +N | 50 | 10.2* | 8.0 | 7.7 | 7.4 | 7.9* |
| +N, nutrients | 0 | 10.2* | 8.0 | 7.5 | 7.3 | 6.9 |
| Rotated Sorghum, C:N ratio | ||||||
| -N | 0 | 8.9 (9.1†) | 7.5 (7.9) | 6.9 (7.3) | 5.7 (6.6) | 5.5(6.3) |
| -N | 25 | 9.0 | 7.2 | 6.5 | 5.8 | 6.1 |
| -N | 50 | 8.9 | 7.1 | 6.6 | 5.7 | 5.7 |
| +N | 0 | 9.1 | 6.9 | 6.7 | 5.8 | 5.9 |
| +N | 25 | 8.9 | 7.3 | 6.9 | 5.9 | 5.8 |
| +N | 50 | 8.4 | 7.1 | 6.5 | 5.8 | 6.3 |
| +N, nutrients | 0 | 8.7 | 7.2 | 6.4 | 6.2 | 6.2 |
| † Values in parentheses are soil C:N ratios prior to study initiation in 2008. * Indicates soil C:N ratios are significantly different at 5% level from initial C:N ratio values in 2008. | ||||||





