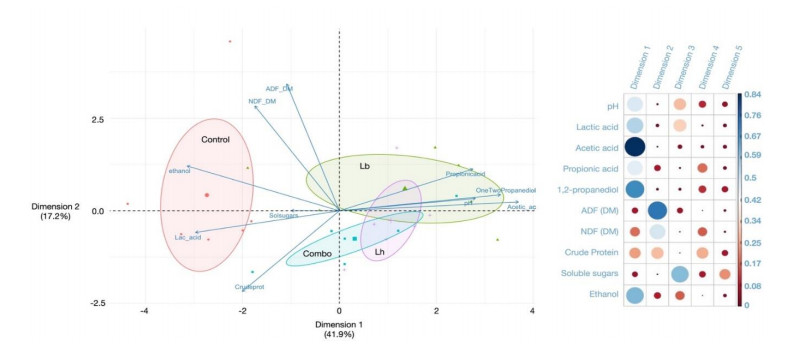
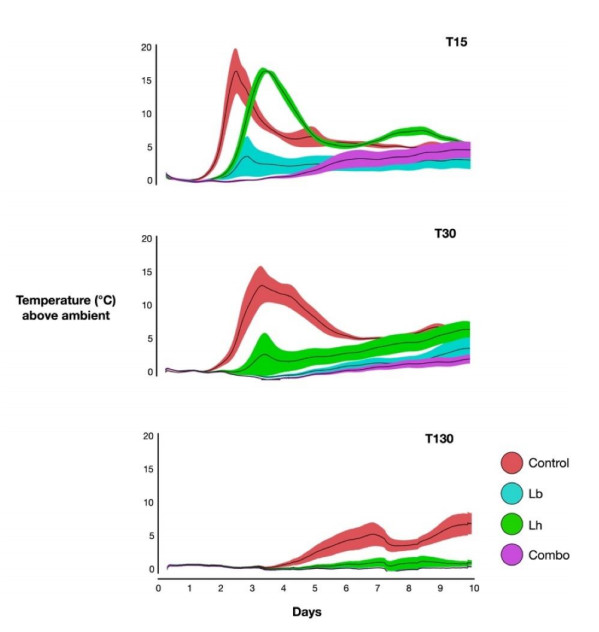
Triticale, a small-grain forage, was harvested for an ensiling experiment evaluating different silage inoculants. Fresh material (mean dry matter 404.1 g kg−1) was wilted, chopped, and assigned to one of four treatment groups including water only (Control), heterofermentative strains Lentilactobacillus buchneri (LB), Lentilactobacillus hilgardii (LH), or combination (Combo) of both. Inoculants were applied at 4.0 x 105 CFU per g of fresh forage, and the Combo contained both strains at 2.0 x 105 CFU per g. Treated forage was packed into 7.57 L mini-silos for openings after 15, 30, and 130 d. Samples were collected at each opening for microbial enumeration of lactic acid bacteria, enterobacteria, yeasts, and moulds. Additional samples were collected for analysis of fermentation profiles, and nutritional analysis following dry matter determination and grinding. Aerobic stability was also evaluated at each opening through a 10-d period. Fermentation profiles were notably affected, including increases in acetic acid (g/kg DM) in LB and Combo treated silage after 15 d which resulted in reduced pH. Aerobic stability was vastly improved through inoculation by prolonging the time to reach 2˚ C above ambient, which was likely a result of decreased yeast counts. Our research validates the use of combined heterofermentative lactic acid bacteria strains on an ensiled small-grain specie, and further agrees with previous findings of prolonged aerobic stability through increases in lactate and acetate in response to co-inoculation.
Citation: Richard Anthony Scuderi, Pascal Drouin, Emmanuelle Apper. Inoculation with heterofermentative strains Lentilactobacillus buchneri CNCM 40788 and Lentilactobacillus hilgardii CNCM I-4785 either alone or combined improves fermentation and aerobic stability of ensiled triticale (X-triticosecale)[J]. AIMS Agriculture and Food, 2023, 8(3): 914-931. doi: 10.3934/agrfood.2023048
Triticale, a small-grain forage, was harvested for an ensiling experiment evaluating different silage inoculants. Fresh material (mean dry matter 404.1 g kg−1) was wilted, chopped, and assigned to one of four treatment groups including water only (Control), heterofermentative strains Lentilactobacillus buchneri (LB), Lentilactobacillus hilgardii (LH), or combination (Combo) of both. Inoculants were applied at 4.0 x 105 CFU per g of fresh forage, and the Combo contained both strains at 2.0 x 105 CFU per g. Treated forage was packed into 7.57 L mini-silos for openings after 15, 30, and 130 d. Samples were collected at each opening for microbial enumeration of lactic acid bacteria, enterobacteria, yeasts, and moulds. Additional samples were collected for analysis of fermentation profiles, and nutritional analysis following dry matter determination and grinding. Aerobic stability was also evaluated at each opening through a 10-d period. Fermentation profiles were notably affected, including increases in acetic acid (g/kg DM) in LB and Combo treated silage after 15 d which resulted in reduced pH. Aerobic stability was vastly improved through inoculation by prolonging the time to reach 2˚ C above ambient, which was likely a result of decreased yeast counts. Our research validates the use of combined heterofermentative lactic acid bacteria strains on an ensiled small-grain specie, and further agrees with previous findings of prolonged aerobic stability through increases in lactate and acetate in response to co-inoculation.

| [1] | Salmon DF, Mergoum M, Gomez-Macpherson H (2004) Triticale production and management. Triticale improvement and production, Rome, Food and Agriculture Organization of the United Nations, 27–32. |
| [2] |
McGoverin CM, Snyders F, Muller N, et al. (2011) A review of triticale uses and the effect of growth environment on grain quality. J Sci Food Agric 91: 1155–1165. https://doi.org/10.1002/jsfa.4338 doi: 10.1002/jsfa.4338

|
| [3] |
Ayalew H, Kumssa TT, Butler TJ, et al. (2018) Triticale improvement for forage and cover crop uses in the Southern Great Plains of the United States. Front Plant Sci 9: 1130. https://doi.org/10.3389/fpls.2018.01130 doi: 10.3389/fpls.2018.01130
|
| [4] |
Muck RE, Nadeau EMG, McAllister TA, et al. (2018) Silage review: Recent advances and future uses of silage additives. J Dairy Sci 101: 3980–4000. https://doi.org/10.3168/jds.2017-13839 doi: 10.3168/jds.2017-13839
|
| [5] |
Driehuis F, Elferink SJWHO, Spoelstra SF (1999) Anaerobic lactic acid degradation during ensilage of whole crop maize inoculated with Lactobacillus buchneri inhibits yeast growth and improves aerobic stability. J Appl Microbiol 87: 583–594. https://doi.org/10.1046/j.1365-2672.1999.00856.x doi: 10.1046/j.1365-2672.1999.00856.x
|
| [6] |
Oude Elferink SJWH, Krooneman J, Gottschal JC, et al. (2001) Anaerobic conversion of lactic acid to acetic acid and 1, 2-propanediol by Lactobacillus buchneri. Appl Environ Microbiol 67: 125–132. https://doi.org/10.1128/AEM.67.1.125-132.2001 doi: 10.1128/AEM.67.1.125-132.2001
|
| [7] |
Zheng J, Wittouck S, Salvetti E, et al. (2020) A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int J Syst Evol Microbiol 70: 2782–2858. https://doi.org/10.1099/ijsem.0.004107 doi: 10.1099/ijsem.0.004107
|
| [8] |
Oliveira AS, Weinberg ZG, Ogunade IM, et al. (2017) Meta-analysis of effects of inoculation with homofermentative and facultative heterofermentative lactic acid bacteria on silage fermentation, aerobic stability, and the performance of dairy cows. J Dairy Sci 100: 4587–4603. https://doi.org/10.3168/jds.2016-11815 doi: 10.3168/jds.2016-11815
|
| [9] |
Kleinschmit DH, Kung L (2006) A meta-analysis of the effects of Lactobacillus buchneri on the fermentation and aerobic stability of corn and grass and small-grain silages. J Dairy Sci 89: 4005–4013. https://doi.org/10.3168/jds.S0022-0302(06)72444-4 doi: 10.3168/jds.S0022-0302(06)72444-4
|
| [10] |
Ferrero F, Piano S, Tabacco E, et al. (2019) Effects of conservation period and Lactobacillus hilgardii inoculum on the fermentation profile and aerobic stability of whole corn and sorghum silages: Effects of Lactobacillus hilgardii on corn silage. J Sci Food Agric 99: 2530–2540. https://doi.org/10.1002/jsfa.9463 doi: 10.1002/jsfa.9463
|
| [11] |
Carvalho BF, Ávila CLS, Miguel MGCP, et al. (2015) Aerobic stability of sugar-cane silage inoculated with tropical strains of lactic acid bacteria. Grass Forage Sci 70: 308–323. https://doi.org/10.1111/gfs.12117 doi: 10.1111/gfs.12117
|
| [12] |
Reis CB, de Oliveira dos Santos A, Carvalho BF, et al. (2018) Wild Lactobacillus hilgardii (CCMA 0170) strain modifies the fermentation profile and aerobic stability of corn silage. J Appl Anim Res 46: 632–638. https://doi.org/10.1080/09712119.2017.1371609 doi: 10.1080/09712119.2017.1371609
|
| [13] |
Silva EB, Smith ML, Savage RM, et al. (2020) Effects of Lactobacillus hilgardii 4785 and Lactobacillus buchneri 40788 on the bacterial community, fermentation and aerobic stability of high‐moisture corn silage. J Appl Microbiol 130: 1481–1493. https://doi.org/10.1111/jam.14892 doi: 10.1111/jam.14892
|
| [14] |
Drouin P, Tremblay J, Chaucheyras-Durand F (2019) Dynamic succession of microbiota during ensiling of whole plant corn following inoculation with Lactobacillus buchneri and Lactobacillus hilgardii Alone or in Combination. Microorganisms 7: 595. https://doi.org/10.3390/microorganisms7120595 doi: 10.3390/microorganisms7120595
|
| [15] |
Drouin P, Tremblay J, Renaud J, et al. (2020) Microbiota succession during aerobic stability of maize silage inoculated with Lentilactobacillus buchneri NCIMB 40788 and Lentilactobacillus hilgardii CNCM‐I‐4785. Microbiol Open 10: e1153. https://doi.org/10.1002/mbo3.1153 doi: 10.1002/mbo3.1153
|
| [16] |
Drouin P, Tremblay J, da Silva ÉB, et al. (2022) Changes to the microbiome of alfalfa during the growing season and after ensiling with Lentilactobacillus buchneri and Lentilactobacillus hilgardii inoculant. J Appl Microbiol 133: 2331–2347. https://doi.org/10.1111/jam.15641 doi: 10.1111/jam.15641
|
| [17] |
Bergen WG, Byrem TM, Grant AL (1991) Ensiling characteristics of whole-crop small grains harvested at milk and dough stages. J Anim Sci 69: 1766. https://doi.org/10.2527/1991.6941766x doi: 10.2527/1991.6941766x
|
| [18] |
Ozduven ML, Kursun Onal Z, Koc F (2009) The effects of bacterial inoculants and/or enzymes on the fermentation, aerobic stability and in vitro dry and organic matter digestibility characteristics of triticale silages. Kafkas Univ Vet Fak Derg 16: 751–756. https://doi.org/10.9775/kvfd.2009.1628 doi: 10.9775/kvfd.2009.1628
|
| [19] |
Keles G, Demirci U (2011) The effect of homofermentative and heterofermentative lactic acid bacteria on conservation characteristics of baled triticale–Hungarian vetch silage and lamb performance. Anim Feed Sci Technol 164: 21–28. https://doi.org/10.1016/j.anifeedsci.2010.11.017 doi: 10.1016/j.anifeedsci.2010.11.017
|
| [20] |
Duniere L, Xu S, Long J, et al. (2017) Bacterial and fungal core microbiomes associated with small grain silages during ensiling and aerobic spoilage. BMC Microbiol 17: 50. https://doi.org/10.1186/s12866-017-0947-0 doi: 10.1186/s12866-017-0947-0
|
| [21] |
Soundharrajan I, Kim D, Kuppusamy P, et al. (2019) Probiotic and Triticale silage fermentation potential of Pediococcus pentosaceus and Lactobacillus brevis and their impacts on pathogenic bacteria. Microorganisms 7: 318. https://doi.org/10.3390/microorganisms7090318 doi: 10.3390/microorganisms7090318
|
| [22] |
Drouin P, Ferrero F (2020) Testing selectivity of bacterial and fungal culture media compared to original silage samples using next generation sequencing. J Microbiol Methods 179: 106088. https://doi.org/10.1016/j.mimet.2020.106088 doi: 10.1016/j.mimet.2020.106088
|
| [23] | Kassambara A, Fabian M (2020) Factoextra: Extract and visualize the results of Mulitvariate Data Analyses. Available from: http://www.sthda.com/english/rpkgs/factoextra. |
| [24] |
Lê S, Josse J, Husson F (2008) FactoMineR: An R package for multivariate analysis. J Stat Soft 25: 1–18. https://doi.org/10.18637/jss.v025.i01 doi: 10.18637/jss.v025.i01
|
| [25] | Wei T S (2021) R Package 'corrplot': Visualization of a correlation matrix. Available from: https://github.com/taiyun/corrplot. |
| [26] |
Arriola KG, Vyas D, Kim D, et al. (2021) Effect of Lactobacillus hilgardii, Lactobacillus buchneri, or their combination on the fermentation and nutritive value of sorghum silage and corn silage. J Dairy Sci 104: 9664–9675. https://doi.org/10.3168/jds.2020-19512 doi: 10.3168/jds.2020-19512
|
| [27] |
Kung L, Shaver RD, Grant RJ, et al. (2018) Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J Dairy Sci 101: 4020–4033. https://doi.org/10.3168/jds.2017-13909 doi: 10.3168/jds.2017-13909
|
| [28] |
Santos WP, Ávila CLS, Pereira MN, et al. (2017) Effect of the inoculation of sugarcane silage with and on feeding behavior and milk yield of dairy cows. J Anim Sci 95: 4613–4622. https://doi.org/10.2527/jas.2017.1526 doi: 10.2527/jas.2017.1526
|
| [29] |
Pedroso A de F, Nussio LG, Loures DRS, et al. (2008) Fermentation, losses, and aerobic stability of sugarcane silages treated with chemical or bacterial additives. Sci Agric (Piracicaba, Braz) 65: 589–594. https://doi.org/10.1590/S0103-90162008000600004 doi: 10.1590/S0103-90162008000600004
|
| [30] | Rooke JA, Hatfield RD (2003) Biochemistry of ensiling. In: Buxton DR, Muck RE, Harrison JH (Eds.), Agronomy Monographs, Madison, WI, USA, American Society of Agronomy, Crop Science Society of America, Soil Science Society of America, 95–139. |
| [31] |
Blajman JE, Páez RB, Vinderola CG, et al. (2018) A meta-analysis on the effectiveness of homofermentative and heterofermentative lactic acid bacteria for corn silage. J Appl Microbiol 125: 1655–1669. https://doi.org/10.1111/jam.14084 doi: 10.1111/jam.14084
|
| [32] |
Blajman JE, Vinderola G, Páez RB, et al. (2020) The role of homofermentative and heterofermentative lactic acid bacteria for alfalfa silage: A meta-analysis. J Agric Sci 158: 107–118. https://doi.org/10.1017/S0021859620000386 doi: 10.1017/S0021859620000386
|
| [33] | Pahlow G, Muck RE, Driehuis F, et al. (2003) Microbiology of ensiling. In: Buxton DR, Muck RE, Harrison JH (Eds.), Agronomy Monographs, Madison, WI, USA, American Society of Agronomy, Crop Science Society of America, Soil Science Society of America, 31–93. |
| [34] |
Driehuis F, Wilkinson JM, Jiang Y, et al. (2018) Silage review: Animal and human health risks from silage. J Dairy Sci 101: 4093–4110. https://doi.org/10.3168/jds.2017-13836 doi: 10.3168/jds.2017-13836
|
| [35] |
Ferrero F, Tabacco E, Borreani G (2021) Lentilactobacillus hilgardii inoculum, dry matter contents at harvest and length of conservation affect fermentation characteristics and aerobic stability of corn silage. Front Microbiol 12: 675563. https://doi.org/10.3389/fmicb.2021.675563 doi: 10.3389/fmicb.2021.675563
|
| [36] |
Ávila CLS, Carvalho BF, Pinto JC, et al. (2014) The use of Lactobacillus species as starter cultures for enhancing the quality of sugar cane silage. J Dairy Sci 97: 940–951. https://doi.org/10.3168/jds.2013-6987 doi: 10.3168/jds.2013-6987
|
| [37] |
Valerio F, Lavermicocca P, Pascale M, et al. (2004) Production of phenyllactic acid by lactic acid bacteria: An approach to the selection of strains contributing to food quality and preservation. FEMS Microbiol Lett 233: 289–295. https://doi.org/10.1111/j.1574-6968.2004.tb09494.x doi: 10.1111/j.1574-6968.2004.tb09494.x
|
| [38] |
Pessione E (2012) Lactic acid bacteria contribution to gut microbiota complexity: Lights and shadows. Front Cell Inf Microbio 2: 00086. https://doi.org/10.3389/fcimb.2012.00086 doi: 10.3389/fcimb.2012.00086
|
| [39] |
Junges D, Schmidt P, Novinski CO, et al. (2013) Additive containing homo and heterolactic bacteria on the fermentation quality of maize silage. Acta Sci Anim Sci 35: 371–377. https://doi.org/10.4025/actascianimsci.v35i4.18833 doi: 10.4025/actascianimsci.v35i4.18833
|
Figures(2) / Tables(4)

Richard Anthony Scuderi, Pascal Drouin, Emmanuelle Apper. Inoculation with heterofermentative strains Lentilactobacillus buchneri CNCM 40788 and Lentilactobacillus hilgardii CNCM I-4785 either alone or combined improves fermentation and aerobic stability of ensiled triticale (X-triticosecale)[J]. AIMS Agriculture and Food, 2023, 8(3): 914-931. doi: 10.3934/agrfood.2023048







 DownLoad:
DownLoad: