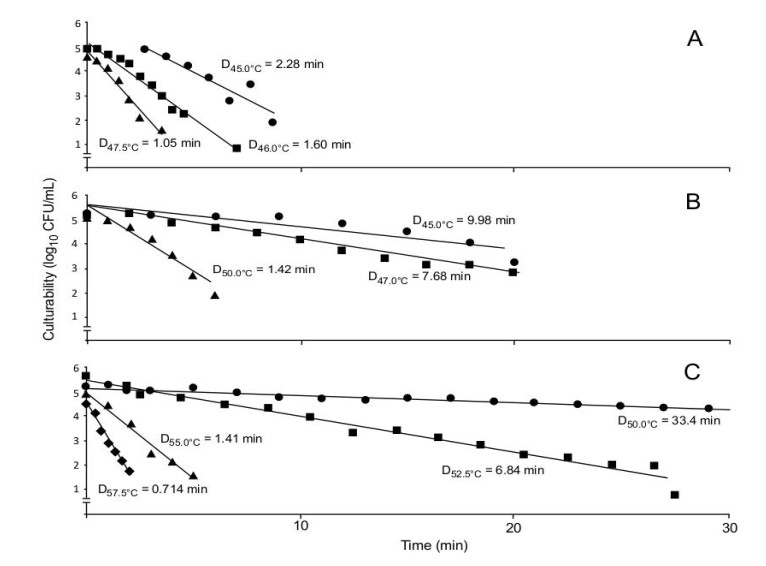
A yeast, isolated from grape juice concentrate by a grape juice processing company in central Washington, was subjected to thermal processing in 52° Brix grape juice concentrate, commercial grape juice, and Yeast-Mold (YM) medium to determine D-values and z-values in the search for an effective means of control. At 50 ℃ in grape juice concentrate the D-value was 33.44 minutes, dropping rapidly as the temperature increased to 52.5 ℃, 55 ℃, 57.5 ℃, and 60 ℃. From this data, a z-value was determined to be 4.38 ℃. Grape juice and YM broth were similarly tested. Commercial grape juice at 45 ℃, 47 ℃, and 50 ℃ showed D-values of 9.80, 7.68, and 1.42, respectively, resulting in a z-value of 5.69 ℃. The isolate heat treated in YM broth at 40 ℃, 45 ℃, 46 ℃, 47.5 ℃, and 50 ℃ showed no effect at 40 ℃ but at higher temperatures, with a D-value of 2.28 at 45 ℃, 1.61 at 46 ℃, 1.05 at 47.5 ℃, and with a z-value of 7.46 ℃. This data suggests that an increase in the concentration of sugars in the environment has a protective effect on the yeast against elevated temperatures. Even so, pasteurization at 90 ℃ for thirty seconds appears to be an effective means of control for the yeast investigated. A yeast with the identical colony and cell morphology was isolated from 52° Brix grape juice concentrate from the same grape juice processing company. Both isolates were subjected to 28S DNA sequencing and FAME analysis for identification and determined to be Zygosaccharomyces rouxii, a known problem yeast in the grape juice and wine-making industry, to this point unknown in the central Washington region.
Citation: Wu-Joo Lee, Barbara Nielsen, Frank L. Younce, Charles G. Edwards, Gülhan Ünlü. Polyphasic identification of a Zygosaccharomyces rouxii isolated from grape juice concentrate and its control using thermal processing[J]. AIMS Agriculture and Food, 2023, 8(2): 662-673. doi: 10.3934/agrfood.2023036
A yeast, isolated from grape juice concentrate by a grape juice processing company in central Washington, was subjected to thermal processing in 52° Brix grape juice concentrate, commercial grape juice, and Yeast-Mold (YM) medium to determine D-values and z-values in the search for an effective means of control. At 50 ℃ in grape juice concentrate the D-value was 33.44 minutes, dropping rapidly as the temperature increased to 52.5 ℃, 55 ℃, 57.5 ℃, and 60 ℃. From this data, a z-value was determined to be 4.38 ℃. Grape juice and YM broth were similarly tested. Commercial grape juice at 45 ℃, 47 ℃, and 50 ℃ showed D-values of 9.80, 7.68, and 1.42, respectively, resulting in a z-value of 5.69 ℃. The isolate heat treated in YM broth at 40 ℃, 45 ℃, 46 ℃, 47.5 ℃, and 50 ℃ showed no effect at 40 ℃ but at higher temperatures, with a D-value of 2.28 at 45 ℃, 1.61 at 46 ℃, 1.05 at 47.5 ℃, and with a z-value of 7.46 ℃. This data suggests that an increase in the concentration of sugars in the environment has a protective effect on the yeast against elevated temperatures. Even so, pasteurization at 90 ℃ for thirty seconds appears to be an effective means of control for the yeast investigated. A yeast with the identical colony and cell morphology was isolated from 52° Brix grape juice concentrate from the same grape juice processing company. Both isolates were subjected to 28S DNA sequencing and FAME analysis for identification and determined to be Zygosaccharomyces rouxii, a known problem yeast in the grape juice and wine-making industry, to this point unknown in the central Washington region.

| [1] |
Rojo M, Arroyo Lopez F, Lerena M, et al. (2014) Effects of pH and sugar concentration in Zygosaccharomyces rouxii growth and time for spoilage in concentrated grape juice at isothermal and non-isothermal conditions. Food Microbiol 38: 143–150. https://doi.org/10.1016/j.fm.2013.08.017 doi: 10.1016/j.fm.2013.08.017

|
| [2] |
Combina M, Daguerre C, Massera A, et al. (2008) Yeast Identification in grape juice concentrates from Argentina. Lett Appl Microbiol 46: 192–197. https://doi.org/10.1111/j.1472-765X.2007.02291.x doi: 10.1111/j.1472-765X.2007.02291.x
|
| [3] | Bates R, Morris J, Crandall P (2001) Principles and Practices of small- and medium- scale fruit juice processing. Food and Agriculture Organization of the United Nations, Rome, Italy. |
| [4] |
Rojo M, Palazzolo C, Cuello R, et al. (2017) Incidence of osmophilic yeasts and Zygosaccharomyces rouxii during the production of concentrate grape juice. Food Microbiol 64: 7–14. https://doi.org/10.1016/j.fm.2016.11.017 doi: 10.1016/j.fm.2016.11.017
|
| [5] |
Barata A, Gonzalez S, Malfeito-Ferreira M, et al. (2008a) Sour rot-damaged grapes are sources of wine spoilage yeasts. FEMS Yeast Res 8: 1008–1017. https://doi.org/10.1111/j.1567-1364.2008.00399.x doi: 10.1111/j.1567-1364.2008.00399.x
|
| [6] |
Barata A, Seborro F, Belloch C, et al. (2008b) Ascomycetous yeast species recovered from grapes damaged by honeydew and sour rot. J Appl Microbiol 104: 1182–1191. https://doi.org/10.1111/j.1365-2672.2007.03631.x doi: 10.1111/j.1365-2672.2007.03631.x
|
| [7] |
Legen J, Voysey P (1991) Yeast spoilage of bakery products and ingredients. J Appl Bacteriol 70: 361–371. https://doi.org/10.1111/j.1365-2672.1991.tb02950.x doi: 10.1111/j.1365-2672.1991.tb02950.x
|
| [8] |
Martorell P, Stratford M, Steele H, et al. (2006) Physiological characterization of spoilage strains of Zygosaccharomyces bailii and Zygosaccharomyces rouxii isolated from high sugar environments. Int J Food Microbiol 114: 234–242. https://doi.org/10.1016/j.ijfoodmicro.2006.09.014 doi: 10.1016/j.ijfoodmicro.2006.09.014
|
| [9] |
Dakal T, Solieri L, Giudici P (2014) Adaptive response and tolorence to sugar and salt stress in the food yeast Zygosaccharomyces rouxii. Int J Food Microbiol 185: 140–157. https://doi.org/10.1016/j.ijfoodmicro.2014.05.015 doi: 10.1016/j.ijfoodmicro.2014.05.015
|
| [10] |
Shearer A, Mazzotta S, Chuyate R, et al. (2002) Heat resistance of juice spoilage microorganisms. J Food Protect 65: 1271–1275. https://doi.org/10.4315/0362-028X-65.8.1271 doi: 10.4315/0362-028X-65.8.1271
|
| [11] |
Corry J (1976) The effect of sugars and polyols on the heat resistance and morphology of osmophilic yeasts. J. Appl Bact 40: 269–276. https://doi.org/10.1111/j.1365-2672.1976.tb04174.x doi: 10.1111/j.1365-2672.1976.tb04174.x
|
| [12] |
Jermini M, Schmidt-Lorenz W (1987) Heat Resistance of vegetative cells and asci of two Zygosaccharomyces yeasts in briths at different water activity values. J Food Protect 50: 835–841. https://doi.org/10.4315/0362-028X-50.10.835 doi: 10.4315/0362-028X-50.10.835
|
| [13] |
Rojo M, Cristani M, Szermanm N, et al. (2019) Reduction of Zygosaccharomyces rouxii population in concentrated grape juices by thermal pasteurization and hydrostatic high pressure processing. Food Bioprocess Tech 12: 781–788. https://doi.org/10.1007/s11947-019-02251-9 doi: 10.1007/s11947-019-02251-9
|
| [14] |
Gilland R (1959) Determination of yeast viability. J Inst Brew 65: 424–429. https://doi.org/10.1002/j.2050-0416.1959.tb01482.x doi: 10.1002/j.2050-0416.1959.tb01482.x
|
| [15] |
Kucsera J, Yarita K, Takeo K (2000) Simple detection method for distinguishing dead and living yeast colonies. J Microbial Meth 41: 19–21. https://doi.org/10.1016/S0167-7012(00)00136-6 doi: 10.1016/S0167-7012(00)00136-6
|
| [16] |
Boynton P, Greig D (2014) The ecology and evolution of non-domesticated Saccharomyces species. Yeast 31: 449–462. https://doi.org/10.1002/yea.3040 doi: 10.1002/yea.3040
|
| [17] |
Molohon K, Melby J, Lee J, et al. (2011) Structure determination and inception of biosynthetic intermediates for the Plantazolicin class of highly discriminating antibiotics. ACS Chem Biol 6: 1307–1313. https://doi.org/10.1021/cb200339d doi: 10.1021/cb200339d
|
| [18] |
Grady E, MacDonald J, Liu L, et al. (2016) Current knowledge and perspectives of Paenibacillus: A review Microb Cell Fact 15: 203. https://doi.org/10.1186/s12934-016-0603-7 doi: 10.1186/s12934-016-0603-7
|
| [19] |
Logan N, De Clerck E, Lebbe L, et al. (2004) Paenibacillus cineris sp. nov. and Paenibacillus cookii sp. nov., from Antarctic volcanic soils and a geletin-processing plant. Int J Syst Evol Micr 54: 1071–1076. https://doi.org/10.1099/ijs.0.02967-0 doi: 10.1099/ijs.0.02967-0
|
| [20] |
Zuehlke J, Petrova B, Edwards C (2013) Advances in the control of wine spoilage by Zygosaccharomyces and Dekkera/Brettanomyces. Annu Rev Food Sci Technol 4: 57–78. https://doi.org/10.1146/annurev-food-030212-182533 doi: 10.1146/annurev-food-030212-182533
|
| [21] |
Membre J, Kubaczka M, and Chene C (1999) Combined effects of pH and sugar on growth rate of Zygosaccharomyces rouxii, a bakery productspoilage yeast. Appl Environ Microbiol 65: 4921–4925. https://doi.org/10.1128/AEM.65.11.4921-4925.1999 doi: 10.1128/AEM.65.11.4921-4925.1999
|
| [22] |
Jordao A, Vilela A, Cosme F (2015) From sugar of grape to alcohol of wine: Sensorial impact of alcohol in wine. Beverages 1: 292–310. https://doi.org/10.3390/beverages1040292 doi: 10.3390/beverages1040292
|
| [23] |
Dong K, Pan H, Yang D, et al. (2020) Induction, detection, formation, and resuscitation of viable but non-culturable state microorganisms. Compr Rev Food Sci Food Saf 19:149–183. https://doi.org/10.1111/1541-4337.12513 doi: 10.1111/1541-4337.12513
|
| [24] |
Navarro Y, Torija M, Mas A, et al. (2020) Viability-PCR allows monitoring yeast population dynamics in mixed fermentations including viable but non-culturable yeasts. Foods 9: 1373. https://doi.org/10.3390/foods9101373 doi: 10.3390/foods9101373
|
| [25] |
Casadei M, Ingram R, Skinner R, et al. (2000) Heat resistance of Paenibacillus polymyxa in relation to pH and acidulants. J Appl Microbio 89: 801–806. https://doi.org/10.1046/j.1365-2672.2000.01181.x doi: 10.1046/j.1365-2672.2000.01181.x
|
| [26] |
Jessen J, Sveinsson T, Scully S, et al. (2015) Ethanol production by a Paenibacillus species isolated form an Iclelandic hot spring—Production yields from complex biomass. Icel Agric Sci 28: 15–24. https://doi.org/10.16886/IAS.2015.02 doi: 10.16886/IAS.2015.02
|


Figures(3)

Wu-Joo Lee, Barbara Nielsen, Frank L. Younce, Charles G. Edwards, Gülhan Ünlü. Polyphasic identification of a Zygosaccharomyces rouxii isolated from grape juice concentrate and its control using thermal processing[J]. AIMS Agriculture and Food, 2023, 8(2): 662-673. doi: 10.3934/agrfood.2023036







 DownLoad:
DownLoad: