Citation: Ludovic Mathys. Valuing tradeability in exponential Lévy models[J]. Quantitative Finance and Economics, 2020, 4(3): 459-488. doi: 10.3934/QFE.2020021

| [1] |
Acharya V, Pedersen L (2005) Asset pricing with liquidity risk. J Financ Econ 77: 375-410. doi: 10.1016/j.jfineco.2004.06.007

|
| [2] |
Amihud Y, Mendelson H (1986) Asset pricing and the bid-ask spread. J Financ Econ 17: 223-249. doi: 10.1016/0304-405X(86)90065-6
|
| [3] |
Amihud Y, Mendelson H (1989) The effects of beta, bid-ask spread, residual risk, and size on stock returns. J Financ 44: 479-486. doi: 10.1111/j.1540-6261.1989.tb05067.x
|
| [4] |
Amihud Y, Mendelson H, Pedersen L (2005) Liquidity and asset prices. Found Trends Financ 1: 269-364. doi: 10.1561/0500000003
|
| [5] | Applebaum D (2009) Lévy processes and stochastic calculus, Cambridge Studies in Advanced Mathematics, Cambridge University Press, Cambridge. |
| [6] |
Battauz A, De Donno M, Sbuelz A (2012) Real options with a double continuation region. Quantit Financ 12: 465-475. doi: 10.1080/14697688.2010.484024
|
| [7] |
Battauz A, De Donno M, Sbuelz A (2015) Real options and American derivatives: The double continuation region. Manage Sci 61: 1094-1107. doi: 10.1287/mnsc.2013.1891
|
| [8] |
Cai N, Kou S (2011) Option pricing under a mixed-exponential jump diffusion model. Manage Sci 57: 2067-2081. doi: 10.1287/mnsc.1110.1393
|
| [9] |
Cai N, Sun L (2014) Valuation of stock loans with jump risk. J Econ Dyn Control 40: 213-241. doi: 10.1016/j.jedc.2014.01.004
|
| [10] |
Carr P (1998) Randomization and the American put. Rev Financ Stud 11: 597-626. doi: 10.1093/rfs/11.3.597
|
| [11] |
Carr P, Madan D (1999) Option valuation using the fast Fourier transform. J Comput Financ 2: 61-73. doi: 10.21314/JCF.1999.043
|
| [12] |
Cetin U, Jarrow R, Protter P (2004) Liquidity risk and arbitrage pricing theory. Financ Stoch 8: 311-341. doi: 10.1007/s00780-004-0123-x
|
| [13] |
Cetin U, Rogers LCG (2007) Modelling liquidity effects in discrete time. Math Financ 17: 15-29. doi: 10.1111/j.1467-9965.2007.00292.x
|
| [14] |
Chesney M, Kempf A (2012) The value of tradeability. Rev Deriv Res 15: 193-216. doi: 10.1007/s11147-012-9074-0
|
| [15] |
Cont R, Voltchkova E (2005) Integro-differential equations for option prices in exponential Lévy models. Financ Stoch 9: 299-325. doi: 10.1007/s00780-005-0153-z
|
| [16] |
De Donno M, Palmowski Z, Tumilewicz J (2020) Double continuation regions for American and swing options with negative discount rate in Lévy models. Math Financ 30: 196-227. doi: 10.1111/mafi.12218
|
| [17] |
Detemple J, Kitapbayev Y (2018) On American VIX options under the generalized 3/2 and 1/2 models. Math Financ 28: 550-581. doi: 10.1111/mafi.12153
|
| [18] |
Fajardo J, Mordecki E (2014) Skewness premium with Lévy processes. Quant Financ 14: 1619-1626. doi: 10.1080/14697688.2011.618809
|
| [19] |
Filipovic D, Kitapbayev Y (2018) On the American swaption in the linear-rational framework. Quant Financ 18: 1865-1876. doi: 10.1080/14697688.2018.1446547
|
| [20] |
Garman M, Kohlhagen S (1983) Foreign currency option values. J Int Money Financ 2: 231-237. doi: 10.1016/S0261-5606(83)80001-1
|
| [21] | Gerber H, Shiu E (1994) Option pricing by Esscher transforms. Trans Society Actuaries 46: 99-191. |
| [22] | Gökay S, Roch A, Soner M (2011) Liquidity models in continuous and discrete time, In Advanced Mathematical Methods for Finance, Springer, Heidelberg, 333-365. |
| [23] |
Harrison M, Pliska S (1981) Martingales and stochastic integrals in the theory of continuous trading. Stoch Processes Appl 11: 215-260. doi: 10.1016/0304-4149(81)90026-0
|
| [24] |
Jarrow R (1994) Derivative securities markets, market manipulation and option pricing theory. J Financ Quant Anal 29: 241-261. doi: 10.2307/2331224
|
| [25] |
Jeanblanc M, Chesney M (2004) Pricing American currency options in an exponential Lévy model. Appl Math Financ 11: 207-225. doi: 10.1080/1350486042000249336
|
| [26] | Jeanblanc M, Yor M, Chesney M (2006) Mathematical methods for financial markets, Springer Finance, Springer, Berlin. |
| [27] |
Koziol C, Sauerbier P (2007) Valuation of bond illiquidity: An option-theoretical approach. J Fixed Income 16: 81-107. doi: 10.3905/jfi.2007.683320
|
| [28] | Kyprianou A (2006) Fluctuations of Lévy processes with applications: Introductory lectures, Universitext, Springer, Berlin. |
| [29] |
Lamberton D, Mikou M (2011) The smooth-fit property in an exponential Lévy model. J Appl Probab 49: 137-149. doi: 10.1017/S0021900200008901
|
| [30] |
Longstaff F (1995) How much can marketability affect securities values. J Financ 50: 1767-1774. doi: 10.1111/j.1540-6261.1995.tb05197.x
|
| [31] |
Longstaff F (2018) Valuing thinly traded assets. Manage Sci 64: 3868-3878. doi: 10.1287/mnsc.2016.2718
|
| [32] | Mathys L (2019) On extensions of the Barone-Adesi & Whaley method to price American-type options. Working paper. Available at SSRN 3482064. |
| [33] |
Merton R (1976) Option pricing when underlying stock returns are discontinuous. J Financ Econ 3: 125-144. doi: 10.1016/0304-405X(76)90022-2
|
| [34] |
Mordecki E (2002) Optimal stopping and perpetual options for Lévy processes. Financ Stoch 6: 473-493. doi: 10.1007/s007800200070
|
| [35] |
McDonald R, Siegel D (1986) The value of waiting to invest. Q J Econ 101: 707-727. doi: 10.2307/1884175
|
| [36] | Peskir G (2007) A Change-of-variable formula with local time on surface. In Séminaire de Probabilités XL, Springer, Berlin, 69-96. |
| [37] | Peskir G, Shiryaev A (2006) Optimal stopping and free-boundary problems, Lectures in Mathematics, Birkhäuser, Zurich. |
| [38] |
Pham H (1997) Optimal stopping, free boundary and American option in a jump-diffusion model. Appl Math Optim 35: 145-164. doi: 10.1007/s002459900042
|
| [39] | Pham H (1998) Optimal stopping of controlled jump diffusion processes: A viscosity solution approach. J Math Syst Estim Control 8: 1-27. |
| [40] | Sato KI (1999) Lévy processes and infinitely divisible distributions, Cambridge Studies in Advanced Mathematics, Cambridge University Press, Cambridge. |
| [41] |
Takahashi A, Yamazaki A (2009) Efficient static replication of European options under exponential Lévy models. J Futures Mark 29: 1-15. doi: 10.1002/fut.20339
|
| [42] |
Trigeorgis L, Tsekrekos A (2018) Real options in operations research: A review. Eur J Oper Res 270: 1-24. doi: 10.1016/j.ejor.2017.11.055
|
| [43] | Voltchkova E (2005) Equations intégro-différentielles d'évolution: Méthodes numériques et applications en finance. PhD thesis, Ecole Polytechnique. |
 QFE-04-03-021-s001.pdf QFE-04-03-021-s001.pdf |
 |
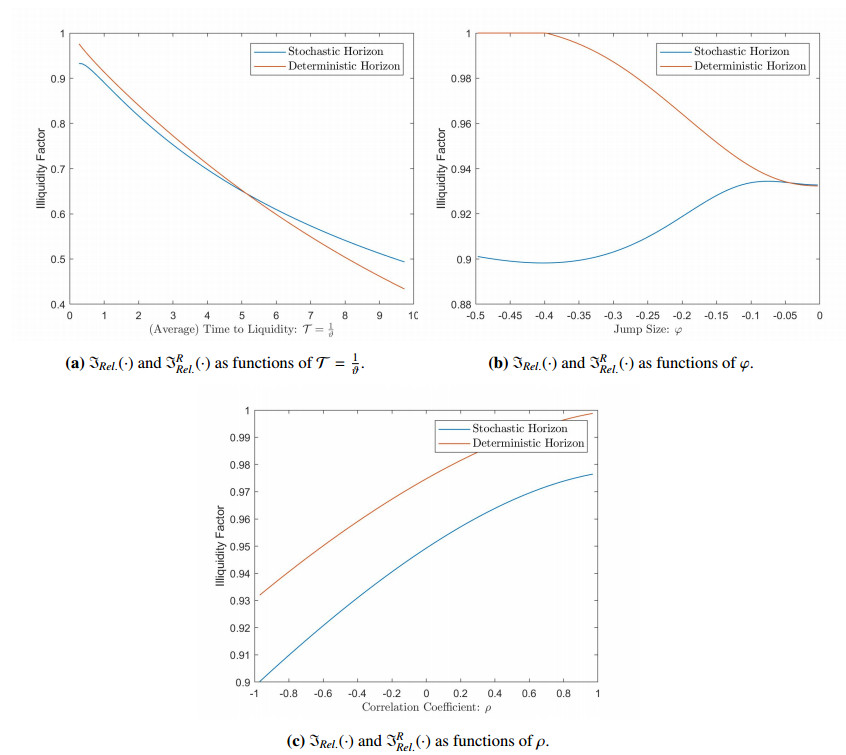
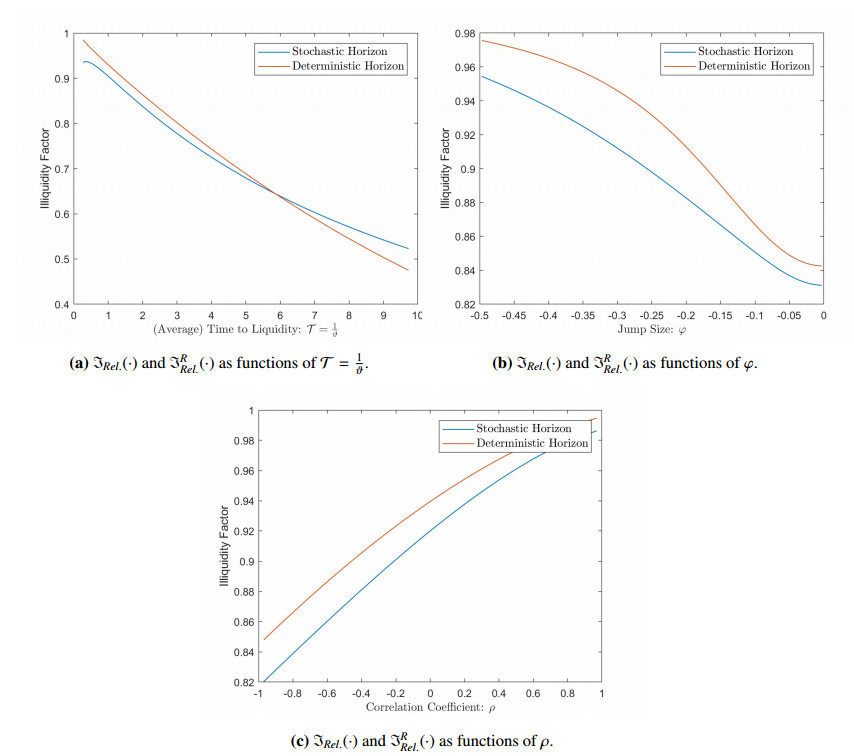
Figures(2) / Tables(4)

Ludovic Mathys. Valuing tradeability in exponential Lévy models[J]. Quantitative Finance and Economics, 2020, 4(3): 459-488. doi: 10.3934/QFE.2020021







 DownLoad:
DownLoad: