Citation: Francisco Les, Zsuzsa Iffiú-Soltész, Josep Mercarder, Christian Carpéné. Tyramine activates lipid accumulation in rat adipocytes: influences of in vitro and in vivo administration[J]. AIMS Molecular Science, 2017, 4(3): 339-351. doi: 10.3934/molsci.2017.3.339

| [1] | Maguire JJ, Parker WA, Foord SM, et al. (2009) International Union of Pharmacology. LXXII. Recommendations for trace amine receptor nomenclature. Pharmacol Rev 61: 1-8. |
| [2] |
Fankhauser C, Charieras T, Caille D, et al. (1994) Interaction of MAO inhibitors and dietary tyramine: a new experimental model in the conscious rat. J Pharmacol Toxicol Methods 32: 219-224. doi: 10.1016/1056-8719(94)90091-4

|
| [3] |
Cantarini MV, Painter CJ, Gilmore EM, et al. (2004) Effect of oral linezolid on the pressor response to intravenous tyramine. Br J Clin Pharmacol 58: 470-475. doi: 10.1111/j.1365-2125.2004.02186.x
|
| [4] |
DeMarcaida JA, Schwid SR, White WB, et al. (2006) Effects of tyramine administration in Parkinson's disease patients treated with selective MAO-B inhibitor rasagiline. Mov Disord 21: 1716-1721. doi: 10.1002/mds.21048
|
| [5] |
Finberg JP, Gillman K (2011) Selective inhibitors of monoamine oxidase type B and the "cheese effect". Int Rev Neurobiol 100: 169-190. doi: 10.1016/B978-0-12-386467-3.00009-1
|
| [6] | Marquet A, Kupas K, Johne A, et al. (2012) The effect of safinamide, a novel drug for Parkinson's disease, on pressor response to oral tyramine: a randomized, double-blind, clinical trial. Clin Pharmacol Ther 92: 450-457. |
| [7] | Gillman PK (2016) Monoamine Oxidase Inhibitors: a review concerning dietary tyramine and drug interactions. PsychoTropical Commentaries 1: 1-90. |
| [8] | Tiller JW, Dowling JT, Tung LH, et al. (1985) Tyramine-induced cardiac arrhythmias. N Engl J Med 313: 266-267. |
| [9] |
Komprda T, Sladkova P, Dohnal V (2009) Biogenic amine content in dry fermented sausages as influenced by a producer, spice mix, starter culture, sausage diameter and time of ripening. Meat Sci 83: 534-542. doi: 10.1016/j.meatsci.2009.07.002
|
| [10] |
Ubaldo JC, Carvalho AF, Fonseca LM, et al. (2015) Bioactive amines in Mozzarella cheese from milk with varying somatic cell counts. Food Chem 178: 229-235. doi: 10.1016/j.foodchem.2015.01.084
|
| [11] |
Rumore MM, Roth M, Orfanos A (2010) Dietary tyramine restriction for hospitalized patients on linezolid: an update. Nutr Clin Pract 25: 265-269. doi: 10.1177/0884533610368711
|
| [12] |
Jansen SC, Van Dusseldorp M, Bottema KC, et al. (2003) Intolerance to dietary biogenic amines: a review. Ann Allergy Asthma Immunol 91: 233-240. doi: 10.1016/S1081-1206(10)63523-5
|
| [13] |
Shen SH, Wertz DL, Klinman JP (2012) Implication for functions of the ectopic adipocyte copper amine oxidase (AOC3) from purified enzyme and cell-based kinetic studies. PLoS One 7: e29270. doi: 10.1371/journal.pone.0029270
|
| [14] |
Visentin V, Prevot D, Marti L, et al. (2003) Inhibition of rat fat cell lipolysis by monoamine oxidase and semicarbazide-sensitive amine oxidase substrates. Eur J Pharmacol 466: 235-243. doi: 10.1016/S0014-2999(03)01562-0
|
| [15] | Marti L, Morin N, Enrique-Tarancon G, et al. (1998) Tyramine and vanadate synergistically stimulate glucose transport in rat adipocytes by amine oxidase-dependent generation of hydrogen peroxide. J Pharmacol Exp Ther 285: 342-349. |
| [16] |
Carpéné C, Daviaud D, Boucher J, et al. (2006) Short- and long-term insulin-like effects of monoamine oxidases and semicarbazide-sensitive amine oxidase substrates in cultured adipocytes. Metabolism 55: 1397-1405. doi: 10.1016/j.metabol.2006.06.011
|
| [17] |
Visentin V, Marq P, Bour S, et al. (2003) Effect of prolonged treatment with tyramine on glucose tolerance in streptozotocin-induced diabetic rats. J Physiol Biochem 59: 225-232. doi: 10.1007/BF03179919
|
| [18] |
Visentin V, Bour S, Boucher J, et al. (2005) Glucose handling in streptozotocin-induced diabetic rats is improved by tyramine but not by the amine oxidase inhibitor semicarbazide. Eur J Pharmacol 522: 139-146. doi: 10.1016/j.ejphar.2005.08.051
|
| [19] |
Lino CdS, Sales TdP, Gomes PB, et al. (2007) Anti-diabetic activity of a fraction from Cissus verticillata and tyramine, its main bioactive constituent, in alloxan-induced diabetic rats. Am J Pharmacol Toxicol 2: 178-188. doi: 10.3844/ajptsp.2007.178.188
|
| [20] |
Carpéné C, Schaak S, Guilbeau-Frugier C, et al. (2016) High intake of dietary tyramine does not deteriorate glucose handling and does not cause adverse cardiovascular effects in mice. J Physiol Biochem 72: 539-553. doi: 10.1007/s13105-015-0456-2
|
| [21] |
Iffiu-Soltesz Z, Prévot D, Carpéné C (2009) Influence of prolonged fasting on monoamine oxidase and semicarbazide-sensitive amine oxidase activities in rat white adipose tissue. J Physiol Biochem 65: 11-23. doi: 10.1007/BF03165965
|
| [22] |
Carpéné C, Galitzky J, Saulnier-Blache JS (2016) Short-term and rapid effects of lysophosphatidic acid on human adipose cell lipolytic and glucose uptake activities. AIMS Molec Sci 3: 222-237. doi: 10.3934/molsci.2016.2.222
|
| [23] |
Atgié C, Sauvant P, Ambid L, et al. (2009) Possible mechanisms of weight loss of Siberian hamsters (Phodopus sungorus sungorus) exposed to short photoperiod. J Physiol Biochem 65: 377-386. doi: 10.1007/BF03185933
|
| [24] |
Fowler CJ, Tipton KF (1981) Concentration dependence of the oxidation of tyramine by the two forms of rat liver mitochondrial monoamine oxidase. Biochem Pharmacol 30: 3329-3332. doi: 10.1016/0006-2952(81)90607-9
|
| [25] |
Carpéné C, Les F, Hasnaoui M, et al. (2016) Anatomical distribution of primary amine oxidase activity in four adipose depots and plasma of severely obese women with or without a dysmetabolic profile. J Physiol Biochem 73: 475-486. doi: 10.1007/s13105-016-0526-0
|
| [26] |
Prévot D, Soltesz Z, Abello V, et al. (2007) Prolonged treatment with aminoguanidine strongly inhibits adipocyte semicarbazide-sensitive amine oxidase and slightly reduces fat deposition in obese Zucker rats. Pharmacol Res 56: 70-79. doi: 10.1016/j.phrs.2007.04.002
|
| [27] | Yraola F, Garcia-Vicente S, Marti L, et al. (2007) Understanding the mechanism of action of the novel SSAO substrate (C7NH10)6(V10O28).2H2O, a prodrug of peroxovanadate insulin mimetics. Chem Biol Drug Des 69: 423-428. |
| [28] |
Gilliam LK, Palmer JP, Taborsky GJ, Jr. (2007) Tyramine-mediated activation of sympathetic nerves inhibits insulin secretion in humans. J Clin Endocrinol Metab 92: 4035-4038. doi: 10.1210/jc.2007-0536
|
| [29] | Carpéné C, Iffiú-Soltész Z (2011) Monoaminergic systems and anti-obesity drug discovery: chronic administration of sympathicomimetic amines, re-uptake inhibitors, or amine oxidase inhibitors? In: Anti-Obesity Drug Discovery and Development (Atta-ur-Rahman & Iqbal Choudhary M Editors) Bentham Sciences Publishers 1: 114-130. |
| [30] |
Suzzi G (2003) Biogenic amines in dry fermented sausages: a review. Int J Food Microbiol 88: 41-54. doi: 10.1016/S0168-1605(03)00080-1
|
| [31] |
Til HP, Falke HE, Prinsen MK, et al. (1997) Acute and subacute toxicity of tyramine, spermidine, spermine, putrescine and cadaverine in rats. Food Chem Toxicol 35: 337-348. doi: 10.1016/S0278-6915(97)00121-X
|
| [32] |
Yonekura T, Kamata S, Wasa M, et al. (1988) Simultaneous determination of plasma phenethylamine, phenylethanolamine, tyramine and octopamine by high-performance liquid chromatography using derivatization with fluorescamine. J Chromatogr 427: 320-325. doi: 10.1016/0378-4347(88)80135-X
|
| [33] |
VanDenBerg CM, Blob LF, Kemper EM, et al. (2003) Tyramine pharmacokinetics and reduced bioavailability with food. J Clin Pharmacol 43: 604-609. doi: 10.1177/0091270003253425
|
| [34] | Korn A, Da Prada M, Raffesberg W, et al. (1988) Tyramine pressor effect in man: studies with moclobemide, a novel, reversible monoamine oxidase inhibitor. J Neural Transm Suppl 26: 57-71. |
| [35] |
McDonald GR, Hudson AL, Dunn SM, et al. (2008) Bioactive contaminants leach from disposable laboratory plasticware. Science 322: 917. doi: 10.1126/science.1162395
|
| [36] |
Mundinger TO, Cummings DE, Taborsky GJ (2006) Direct stimulation of ghrelin secretion by sympathetic nerves. Endocrinology 147: 2893-2901. doi: 10.1210/en.2005-1182
|
| [37] |
Adams F, Boschmann M, Schaller K, et al. (2006) Tyramine in the assessment of regional adrenergic function. Biochem Pharmacol 72: 1724-1729. doi: 10.1016/j.bcp.2006.09.004
|
| [38] |
Morin N, Visentin V, Calise D, et al. (2002) Tyramine stimulates glucose uptake in insulin-sensitive tissues in vitro and in vivo via its oxidation by amine oxidases. J Pharmacol Exp Ther 303: 1238-1247. doi: 10.1124/jpet.102.040592
|
| [39] |
Bairras C, Ferrand C, Atgié C (2003) Effect of tyramine, a dietary amine, on glycerol and lactate release by isolated adipocytes from old rats. J Physiol Biochem 59: 161-167. doi: 10.1007/BF03179911
|
| [40] |
Stohs SJ, Hartman MJ (2015) A review of the receptor binding and pharmacological effects of N-methyltyramine. Phytother Res 29: 14-16. doi: 10.1002/ptr.5231
|
| [41] |
Hamasaki H (2016) Functional foods for type 2 diabetes. AIMS Med Sci 3: 278-297. doi: 10.3934/medsci.2016.3.278
|
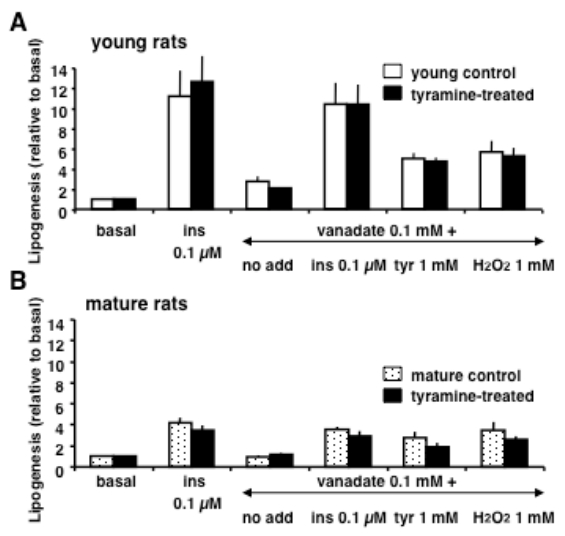
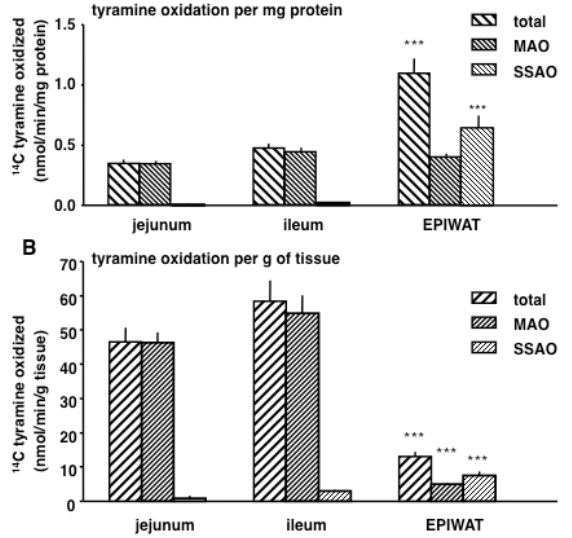
Figures(2) / Tables(2)

Francisco Les, Zsuzsa Iffiú-Soltész, Josep Mercarder, Christian Carpéné. Tyramine activates lipid accumulation in rat adipocytes: influences of in vitro and in vivo administration[J]. AIMS Molecular Science, 2017, 4(3): 339-351. doi: 10.3934/molsci.2017.3.339







 DownLoad:
DownLoad: