Citation: Olaya Muñoz-Azcarate, Ana M González, Marta Santalla. Natural rhizobial diversity helps to reveal genes and QTLs associated with biological nitrogen fixation in common bean[J]. AIMS Microbiology, 2017, 3(3): 435-466. doi: 10.3934/microbiol.2017.3.435

| [1] |
Grange L, Hungria M, Graham PH, et al. (2007) New insights in the origins and evolution of rhizobia that nodulate common bean (Phaseolus vulgaris) in Brazil. Soil Biol Biochem 39: 867–876. doi: 10.1016/j.soilbio.2006.10.008

|
| [2] | CGIAR (Consultative Group on International Agricultural Research), Common Bean, 2012. Available from: http://www.cgiar.org/our-research/crops-factsheets /beans/. |
| [3] |
Aguilar OM, Lopez M, Riccillo PM (2001) The diversity of rhizobia nodulating beans in Northwest Argentina as a source of more efficient inoculant strains. J Biotechnol 91: 181–188. doi: 10.1016/S0168-1656(01)00336-4
|
| [4] | Rodiño AP, Santalla M, De Ron AM, et al. (2010) Co-evolution and migration of bean and rhizobia in Europe, In: Lichtfouse E, Editor, Sociology, organic farming, climate change and soil science, Dordrecht: Springer, 171–188. |
| [5] | Gepts P (1990) Biochemical evidence bearing on the domestication of Phaseolus beans. Econ Bot 44: 28–38. |
| [6] | Papa R, Nanni L, Sicard D, et al. (2006) The evolution of genetic diversity in Phaseolus vulgaris L, In: Darwin's harvest: New approaches to the origins, evolution and conservation of crops, USA: Columbia University Press. |
| [7] | Gepts P, Debouck D (1991) Origin, domestication and evolution of the common bean (Phaseolus vulgaris L.), In: van Schoonhoven A, Voysest O, Editors, Common Beans: Research for Crop Improvement, Wallingford, UK and CIAT, Colombia: CAB Int., 7–53. |
| [8] |
Santalla M, Rodiño AP, De Ron AM (2002) Allozyme evidence supporting Southwestern Europe as a secondary center of genetic diversity for the common bean. Theor Appl Genet 104: 934–944. doi: 10.1007/s00122-001-0844-6
|
| [9] | Gioia T, Logozzo G, Attene G, et al. (2013) Evidence for introduction bottleneck and extensive inter-gene pool (Mesoamerica × Andes) hybridization in the European common bean (Phaseolus vulgaris L.) Germplasm. PLoS One 8: e75974. |
| [10] | Zeven AC (1997) The introduction of common bean (Phaseolus vulgaris L.) into Western Europe and the phenotypic variation of dry beans collected in the Netherlands in 1946. Euphytica 94: 319–328. |
| [11] |
Oldroyd GED (2013) Speak, friend, and enter: signalling systems that promote beneficial symbiotic associations in plants. Nat Rev Microbiol 11: 252–263. doi: 10.1038/nrmicro2990
|
| [12] |
Olivares J, Bedmar EJ, Sanjuan J (2013) Biological nitrogen fixation in the context of global change. Mol Plant Microbe In 26: 486–494. doi: 10.1094/MPMI-12-12-0293-CR
|
| [13] |
Galloway JN, Dentener FJ, Capone DG, et al. (2004) Nitrogen cycles: Past, present and future. Biogeochemistry 70: 153–226. doi: 10.1007/s10533-004-0370-0
|
| [14] |
Bliss FA (1993) Breeding common bean for improved biological nitrogen fixation. Plant Soil 152: 71–70. doi: 10.1007/BF00016334
|
| [15] |
Fageria NK, Melo LC, Ferreira EPB, et al. (2014) Dry matter, grain yield, and yield components of dry bean as influenced by nitrogen fertilization and rhizobia. Commun Soil Sci Plant Anal 45: 111–125. doi: 10.1080/00103624.2013.848877
|
| [16] | Hardarson G (1994) International FAO/IAEA programs on biological nitrogen fixation, In: Graham PH, Sadowsky MJ, Vance CP, Editors, Symbiotic Nitrogen Fixation, Dordrecht: Kluwer Academic Publishers, 189–202. |
| [17] |
Graham PH (1981) Some problems of nodulation and symbiotic nitrogen fixation in Phaseolus vulgaris: a review. Field Crops Res 4: 93–112. doi: 10.1016/0378-4290(81)90060-5
|
| [18] |
Piha MI, Munns DN (1987) Nitrogen-Fixation potential of beans (Phaseolus vulgaris L) compared with other grain legumes under controlled conditions. Plant Soil 98: 169–182. doi: 10.1007/BF02374821
|
| [19] | Wolyn DJ, St Clair DA, DuBois J, et al. (1991) Distribution of nitrogen in common bean (Phaseolus vulgaris L.) genotypes selected for differences in nitrogen fixation ability. Plant Soil 138: 303–311. |
| [20] |
Martínez-Romero E (2003) Diversity of Rhizobium-Phaseolus vulgaris symbiosis: overview and perspectives. Plant Soil 252: 11–23. doi: 10.1023/A:1024199013926
|
| [21] |
Miranda BD, Bliss FA (1991) Selection for increased seed nitrogen accumulation in common bean-implications for improving dinitrogen fixation and seed yield. Plant Breeding 106: 301–311. doi: 10.1111/j.1439-0523.1991.tb00515.x
|
| [22] | Bonilla I, Bolaños L (2010) Mineral nutrition for Legume-Rhizobia Symbiosis: B, Ca, N, P, S, K, Fe, Mo, Co, and Ni: A Review, In: Organic Farming, Pest Control and Remediation of Soil Pollutants, 253–274. |
| [23] |
Divito GA, Sadras VO (2014) How do phosphorous, potassium and sulphur affect plant growth and biological nitrogen fixation in crop and pasture legumes? Field Crop Res 156: 161–171. doi: 10.1016/j.fcr.2013.11.004
|
| [24] | Muñoz-Azcárate O (2014) Sulphur metabolism in the pea-Rhizobium symbiosis, PhD Thesis Dissertation, Public University of Navarre, Spain. |
| [25] | Arrese-Igor C, Gonzalez EM, Marino D, et al. (2011) Physiological responses of legume nodules to drought, In: Anjum NA, Lopez-Lauri F, Editors, Plant Nutrition and Abiotic Stress Tolerance III, Plant Stress, 24–31. |
| [26] |
Aranjuelo I, Arrese-Igor C, Molero G (2014) Nodule performance within a changing environmental context. J Plant Physiol 171: 1076–1090. doi: 10.1016/j.jplph.2014.04.002
|
| [27] |
Bliss FA, Pereira PAA, Araujo RS (1989) Registration of 5 high nitrogen-fixing common bean germplasm lines. Crop Sci 29: 240–241. doi: 10.2135/cropsci1989.0011183X002900010065x
|
| [28] |
De Souza EM, Bassani VL, Sperotto RA, et al. (2016) Inoculation of new rhizobial isolates improves nutrient uptake and growth of bean (Phaseolus vulgaris) and arugula (Eruca sativa). J Sci Food Agric 96: 3446–3453. doi: 10.1002/jsfa.7527
|
| [29] | Sampaio FB, Knupp AM, Fernandes EP, et al. (2016) Morphophysiological characterization of rhizobia isolated from wild genotypes of the common bean. Bioscience J 32: 1502–1511. |
| [30] |
Young JPW, Haukka K (1996) Diversity and phylogeny of rhizobia. New Phytol 133: 87–94. doi: 10.1111/j.1469-8137.1996.tb04344.x
|
| [31] |
Van Berkum P, Terefework Z, Paulin L, et al. (2003) Discordant phylogenies within the rrn loci of Rhizobia.J Bacteriol 185: 2988–2998. doi: 10.1128/JB.185.10.2988-2998.2003
|
| [32] | Moussavi SA, Willems A, Nesme X, et al. (2015) Revised phylogeny of Rhizobiaceae: Proposal of the delineation of Pararhizobium gen. nov., and 13 new species combinations. Syst Appl Microbiol 38: 84–90. |
| [33] | Lindström K, Aserse AA, Mousavi SA, et al. (2013) Taxonomy and evolution of nitrogen-fixing organisms, In: de Bruijn FJ, Editor, Biological nitrogen fixation, John Wiley & Sons, Inc. |
| [34] | Young JPW (2003) The genus name Ensifer Casida 1982 takes priority over Sinorhizobium Chen et al. 1988, and Sinorhizobium morelense Wang et al. 2002 is a later synonym of Ensifer adhaerens Casida 1982. Is the combination 'Sinorhizobium adhaerens' (Casida 1982 ) Willems et al. 2003 legitimate? Request for an Opinion. Int J Syst Evol Microbiol 53: 2107–2110. |
| [35] | CSP-Subcommittee on the taxonomy of Rhizobium and Agrobacterium-diversity, phylogeny and systematics. Available from: http://edzna.ccg.unam.mx/rhizobial-taxonomy/. |
| [36] | Weir BS (2012) The current taxonomy of rhizobia. NZ Rhizobia website. Available from: http://www.rhizobia.co.nz/taxonomy/rhizobia. |
| [37] | Valverde A, Igual JM, Peix A, et al. (2006) R. lusitanum sp. Nov a bacterium that nodulates Phaseolus vulgaris. Int J Syst Evol Microbiol 56: 2631–2637. |
| [38] |
Martínez-Romero E, Segovia L, Mercante FM, et al. (1991) Rhizobium tropici, a novel species nodulating Phaseolus vulgaris L. beans and Leucaena sp tres. Int J Syst Bacteriol 41: 417–426. doi: 10.1099/00207713-41-3-417
|
| [39] |
Segovia L, Young JPW, Martínez-Romero E (1993) Reclassification of American Rhizobium leguminosarum biovar phaseoli type I strains as Rhizobium etli sp. Nov. Int J Syst Bacteriol 43: 374–377. doi: 10.1099/00207713-43-2-374
|
| [40] |
Amarguer N, Macheret V, Laguerre G (1997) Rhizobium gallicum sp. Nov., from Phaseolus vulgaris nodules. Int J Syst Bacteriol 47: 996–1006. doi: 10.1099/00207713-47-4-996
|
| [41] | Ribeiro RA, Martins TB, Ormeño-Orrillo E, et al. (2015) Rhizobium ecuadorense sp. nov., an indigenous N2-fixing symbiont of the Ecuadorian common bean (Phaseolus vulgaris L.) genetic pool. Int J Syst Evol Microbiol 65: 3162–3129. |
| [42] |
Ribeiro RA, Rogel MA, López-López A, et al. (2012) Reclassification of Rhizobium tropici type A strains as Rhizobium leucaenae sp. nov. Int J Syst Evol Microbiol 62: 1179–1184. doi: 10.1099/ijs.0.032912-0
|
| [43] | Jordan DC (1984) Family III Rhizobiaceae, In: Krieg NR, Holt JG, Editors, Bergey's Manual of Systematic Bacteriology, Williams and Wilkins, Baltimore, 234–242. |
| [44] | Lopez-Lopez A, Rogel-Hernandez MA, Barois I, et al. (2012) Rhizobium grahamii sp. nov., from nodules of Dalea leporina, Leucaena leucocephala and Clitoria ternatea, and Rhizobium mesoamericanum sp. nov., from nodules of Phaseolus vulgaris, siratro, cowpea and Mimosa pudica. Int J Syst Evol Microbiol 62: 2264–2271. |
| [45] | Peng GX, Yuan QH, Li HX, et al. (2008) Rhizobium oryzae sp. nov., isolated from the wild rice Oryza alta. Int J Syst Evol Microbiol 58: 2158–2163. |
| [46] | Dall'Agnol RF, Ribeiro RA, Ormeno-Orrillo E, et al. (2013) Rhizobium freirei sp. nov., a symbiont of Phaseolus vulgaris that is very effective at fixing nitrogen. Int J Syst Evol Microbiol 63: 4167–4173. |
| [47] | Mnasri B, Liu TY, Saidi S, et al. (2014) Rhizobium azibense sp. nov., a nitrogen fixing bacterium isolated from root nodules of Phaseolus vulgaris. Int J Syst Evol Microbiol 64: 1501–1506. |
| [48] | Román-Ponce B, Zhang JY, Vásquez-Murrieta MS, et al. (2016) Rhizobium acidisoli sp. nov., isolated from root nodules of Phaseolus vulgaris in acid soils. Int J Syst Evol Microbiol 66: 398–406. |
| [49] | Yan J, Yan H, Liu LX, et al. (2017) Rhizobium hidalgonense sp. nov., a nodule endophitic bacterium of Phaseolus vulgaris in acid soil. Arch Microbiol 199: 97–104. |
| [50] |
Mnasri B, Aouani ME, Mhamdi R (2007) Nodulation and growth of common bean (Phaseolus vulgaris) under water deficiency. Soil Biol Biochem 39: 1744–1750. doi: 10.1016/j.soilbio.2007.01.030
|
| [51] |
Zurdo-Piñeiro JL, García-Fraile P, Rivas, et al. (2009) Rhizobia from Lanzarote, the Canary Islands, that nodulate Phaseolus vulgaris have characteristics in common with Sinorhizobium meliloti isolates from mainland Spain. Appl Environ Microbiol 75: 2354–2359. doi: 10.1128/AEM.02811-08
|
| [52] |
Mhamdi R, Laguerre G, Aouani ME, et al. (2002) Different species and symbiotic genotypes of field rhizobia can nodulate Phaseolus vulgaris in Tunisian soils. FEMS Microbiol Ecol 41: 77–84. doi: 10.1111/j.1574-6941.2002.tb00968.x
|
| [53] | Toledo I, Lloret L, Martínez-Romero E (2003) Sinorhizobium americanus sp. nov., a new Sinorhizobium species nodulating native Acacia spp. in Mexico. Syst Appl Microbiol 26: 54–64. |
| [54] |
Talbi C, Delgado MJ, Girard L, et al. (2010) Burkholderia phymatum strains capable of nodulating Phaseolus vulgaris are present in Moroccan soils. Appl Environ Microbiol 76: 4587–4591. doi: 10.1128/AEM.02886-09
|
| [55] |
Aserse AA, Rasanen LA, Assefa F (2012) Phylogeny and genetic diversity of native rhizobia nodulating common bean (Phaseolus vulgaris L.) in Ethiopia. Syst Appl Microbiol 35: 120–131. doi: 10.1016/j.syapm.2011.11.005
|
| [56] |
Martínez-Romero E, Caballero-Mellado J (1996) Rhizobium polygenies and bacterial genetic diversity. Crit Rev Plant Sci 15: 113–140. doi: 10.1080/07352689.1996.10393183
|
| [57] |
Herrera-Cervera JA, Caballero-Mellado J, Laguerre G, et al. (1999) At least five rhizobial species nodulate Phaseolus vulgaris in a Spanish soil. FEMS Microbiol Ecol 30: 87–97. doi: 10.1111/j.1574-6941.1999.tb00638.x
|
| [58] | Sessitsch A, Hardarson G, Akkermans ADL (1997) Characterization of Rhizobium etli and other Rhizobium spp. that nodulate Phaseolus vulgaris L. in an Austrian soil. Mol Ecol 6: 601–608. |
| [59] | Tjaholeksono A (1993) Caractérisation et diversité des souches de Rhizobium nodulant le haricot (Phaseolus vulgaris L.) cultivé en 3 sites tropicaux, Thesis dissertation, University of Claude Bernard-Lyon. |
| [60] |
Diouf A, de Lajudie P, Neyra M (2000) Polyphasic characterization of rhizobia that nodulate Phaseolus vulgaris in West Africa (Senegal and Gambia). Int J Syst Evol Microbiol 50: 159–170. doi: 10.1099/00207713-50-1-159
|
| [61] |
Beyene D, Kassa S, Ampy F, et al. (2004) Ethiopian soils harbor natural populations of rhizobia that form symbioses with common bean (Phaseolus vulgaris L.). Arch Microbiol 181: 129–136. doi: 10.1007/s00203-003-0636-2
|
| [62] |
Wang H, Man CX, Wang ET, et al. (2009) Diversity of rhizobia and interactions among the host legumes and rhizobial genotypes in an agricultural forestry ecosystem. Plant Soil 314: 169–182. doi: 10.1007/s11104-008-9716-z
|
| [63] |
Lopez-Guerrero MG, Ormeño-Orrillo E, Velázquez E, et al. (2012) Rhizobium etli taxonomy revised with novel genomic data and analyses. Syst Appl Microbiol 35: 353–358. doi: 10.1016/j.syapm.2012.06.009
|
| [64] | Ribeiro RA, Ormeño-Orrillo E, Dall'Agnol RF, et al. (2013) Novel Rhizobium lineages isolated from root nodules of the common bean (Phaseolus vulgaris L.) in Andean and Mesoamerican areas. Res Microbiol 164: 740–748. |
| [65] |
Mostasso FL, Dias BG, Vargas MAT, et al. (2002) Selection of bean (Phaseolus vulgaris L) rhizobial strains for the Brazilian Cerrados. Field Crops Res 73: 121–132. doi: 10.1016/S0378-4290(01)00186-1
|
| [66] |
Hungria M, Campo RJ, Mendes IC (2003) Benefits of inoculation of common bean (Phaseolus vulgaris) crop with efficienty and competitive Rhizobium tropici strains. Biol Fert Soils 39: 88–93. doi: 10.1007/s00374-003-0682-6
|
| [67] |
Martínez-Romero E, Hernández-Lucas I, Peña-Cabriales JJ (1998) Symbiotic performance of some modified Rhizobium etli strains in assays with Phaseolus vulgaris beans that have a high capacity to fix N2. Plant Soil 204: 89–94. doi: 10.1023/A:1004383028331
|
| [68] |
Geniaux E, Flores M, Palacios R, et al. (1995) Presence of megaplasmids in Rhizobium tropici and further evidence of differences between the two R. tropici subtypes. Int J Syst Bacteriol 45: 392–394. doi: 10.1099/00207713-45-2-392
|
| [69] |
Hungria M, Andrade DD, Chueire LM (2000) Isolation and characterization of new efficient and competitive bean (Phaseolus vulgaris L.) rhizobia from Brazil. Soil Biol Biochem 32: 1515–1528. doi: 10.1016/S0038-0717(00)00063-8
|
| [70] | Ormeño-Orrillo E, Menna P, Almeida LG, et al. (2012) Genomic basis of broad host range and environmental adaptability of Rhizobium tropici CIAT899 and Rhizobium sp. PRF 81 which are used in inoculants for common bean (Phaseolus vulgaris L.). BMC Genomics 13: 735. |
| [71] | Ormeño-Orrillo E, Gomes DF, Del Cerro P, et al. (2016) Genome of Rhizobium leucaenae strains CFN 299(T) and CPAO 29.8: searching for genes related to a successful symbiotic performance under stressful conditions. BMC Genomics 17: 534. |
| [72] |
Silva C, Vinuesa P, Eguiarte LE, et al. (2003) Rhizobium etli and Rhizobium gallicum nodulate common bean (Phaseolus vulgaris) in a traditionally managed milpa plot in Mexico: Population genetics and biogeographic implications. Appl Environ Microbiol 69: 884–893. doi: 10.1128/AEM.69.2.884-893.2003
|
| [73] |
Verástegui-Valdés MM, Zhang YJ, Rivera-Orduña FN, et al. (2014) Microsymbionts of Phaseolus vulgaris in acid and alkaline soils of Mexico. Syst Appl Microbiol 37: 605–612. doi: 10.1016/j.syapm.2014.08.005
|
| [74] | Bernal G, Graham PH (2001) Diversity in the rhizobia associated with Phaseolus vulgaris L. in Ecuador, and comparisons with Mexican bean rhizobia. Can J Microbiol 47: 526–534. |
| [75] |
Miranda-Sánchez F, Rivera J, Vinuesa P (2016) Diversity patterns of Rhizobiaceae communities inhabiting soils, root surfaces and nodules reveal a strong selection of rhizobial partners by legumes. Environ Microbiol 18: 2375–2391. doi: 10.1111/1462-2920.13061
|
| [76] | Bitocchi E, Bellucci E, Giardini A, et al. (2013) Molecular analysis of the parallel domestication of the common bean (Phaseolus vulgaris L.) in Mesoamerica and the Andes. New Phytol 197: 300–313. |
| [77] | Valverde G, Otabbong E (1997) Evaluation of N2-fixation measured by the 15N-dilution and N-difference methods in Nicaraguan and Ecuadorian Phaseolus vulgaris L. plants inoculated with Rhizobium leguminosarum biovar. Acta Agr Scand B-S P 47: 71–80. |
| [78] | Kipe-Nolt JA, Montealegre CM, Tohme J (1994) Restriction of nodulation by the broad host range Rhizobium tropici strain CIAT899 in wild accessions of Phaseolus vulgaris L. New Phytol 120: 489–494. |
| [79] | Eardly BD, Wang F, Whittham TS, et al. (1995) Species limits in Rhizobium populations that nodulate the common bean (Phaseolus vulgaris). Appl Environ Microbiol 61: 507–512. |
| [80] | Aguilar OM, Lopez M, Riccillo PM, et al. (1998) Prevalence of the Rhizobium etli-like allele in genes coding for 16S rRNA among the indigenous populations associated with wild beans from the Southern Andes in Argentina. Appl Environ Microbiol 64: 3520–3524. |
| [81] | Aguilar OM, Riva O, Peltzer E (2004) Analysis of R. etli and of its symbiosis with wild Phaseolus vulgaris supports coevolution in centers of host diversification. Proc Natl Acad Sci USA 101: 13548–13553. |
| [82] | Grange L, Hungria M (2004) Genetic diversity of indigenous common bean (Phaseolus vulgaris L.) rhizobia in two Brazilian ecosystems. Soil Biol Biochem 36: 1389–1398. |
| [83] |
Cardoso AA, Andraus MP, Oliveira TC, et al. (2017) Characterization of rhizobia isolates obtained from nodules of wild genotypes of common bean. Braz J Microbiol 48: 43–50. doi: 10.1016/j.bjm.2016.09.002
|
| [84] | Dall'Agnol RF, Ribeiro RA, Delamuta JR, et al. (2014) Rhizobium paranaense sp. nov., an effective N2-fixing symbiont of common bean (Phaseolus vulgaris L.) with broad geographical distribution in Brazil. Int J Syst Evol Microbiol 64: 3222–3229. |
| [85] |
Díaz-Alcántara CA, Ramírez-Bahena MH, Mulas D, et al. (2014) Analysis of rhizobial strains nodulating Phaseolus vulgaris from Hispaniola Island, a geographic bridge between Meso and South America and the first historical link with Europe. Syst Appl Microbiol 37: 149–156. doi: 10.1016/j.syapm.2013.09.005
|
| [86] | Sánchez AC, Gutiérrez RT, Santana RC, et al. (2014) Effects of co-inoculation of native Rhizobium and Pseudomonas strains on growth parameters and yield of two contrasting Phaseolus vulgaris L. genotypes under Cuban soil conditions. Eur J Soil Biol 62: 105–112. |
| [87] | Urzúa H (2005) Beneficios de la fijación simbiótica de nitrógeno en Chile. Cien Inv Agr 32: 133–150. |
| [88] |
Baginsky C, Brito B, Scherson R (2015) Genetic diversity of Rhizobium from nodulating beans grown in a variety of Mediterranean climate soils of Chile. Arch Microbiol 197: 419–429. doi: 10.1007/s00203-014-1067-y
|
| [89] |
Bernal GR, Tlusty B, Estévez de Jensen C, et al. (2004) Characteristics of rhizobia nodulating beans in the central region of Minnesota. Can J Microbiol 50: 1023–1031. doi: 10.1139/w04-092
|
| [90] |
Perez-Ramirez NO, Rogel MA, Wang E, et al. (1998) Seeds of Phaseolus vulgaris bean carry Rhizobium etli. FEMS Microbiol Ecol 26: 289–296. doi: 10.1111/j.1574-6941.1998.tb00513.x
|
| [91] | Martínez E, Pardo MA, Palacios R, et al. (1985) Reiteration of nitrogen-fixation gene-sequences and specificity of Rhizobium in nodulation and nitrogen-fixation in Phaseolus vulgaris. J Gen Microbiol 131: 1779–1786. |
| [92] |
Michiels J, Dombrecht B, Vermeiren N, et al. (1998) Phaseolus vulgaris is a non-selective host for nodulation. FEMS Microbiol Ecol 26: 193–205. doi: 10.1111/j.1574-6941.1998.tb00505.x
|
| [93] |
Valverde A, Velázquez E, Cervantes E, et al. (2011) Evidence of an American origin for Symbiosis-related genes in Rhizobum lusitanum. Appl Environ Microbiol 77: 5665–5670. doi: 10.1128/AEM.02017-10
|
| [94] |
Velázquez A, Martínez-Romero E, Rodriguez-Navarro DN, et al. (2001) Characterization of rhizobial isolates of Phaseolus vulgaris by staircase electrophoresis of low-molecular-weight RNA. Appl Environ Microbiol 67: 1008–1010. doi: 10.1128/AEM.67.2.1008-1010.2001
|
| [95] |
García-Fraile P, Mulas D, Peix A, et al. (2010) Phaseolus vulgaris is nodulated in Northern Spain by Rhizobium leguminosarum strains harboring two nodC alleles present in American Rhizobium etli strains: biogeographical and evolutionary implications. Can J Microbiol 56: 657–666. doi: 10.1139/W10-048
|
| [96] |
Mulas D, García-Fraile P, Carro L, et al. (2011) Distribution and efficiency of Rhizobium leguminosarum strains nodulating Phaseolus vulgaris in Northern Spanish soils: Selection of native strains that replace conventional N fertilization. Soil Biol Biochem 43: 2283–2293. doi: 10.1016/j.soilbio.2011.07.018
|
| [97] | Pohajda I, Babic KH, Rajnovic I, et al. (2016) Genetic diversity and symbiotic efficiency of indigenous common bean rhizobia in Croatia. Food Technol Biotech 54: 468–474. |
| [98] | Boumouch I, Brhada F, Filali-Maltouf A (2001) Selection of osmotolerant and effective strains of Rhizobium for inoculation of common bean (Phaseolus vulgaris L.) in Moroccan saline soils. Agronomie 21: 591–599. |
| [99] |
Priefer UB, Aurag J, Boesten B, et al. (2001) Characterisation of Phaseolus symbionts isolated from Mediterranean soils and analysis of genetic factors related to pH tolerance. J Biotechnol 91: 223–236. doi: 10.1016/S0168-1656(01)00329-7
|
| [100] |
Faghire M, Mandri B, Oufdou K, et al. (2012) Identification at the species and symbiovar levels of strains nodulating Phaseous vulgaris in saline soils of the Marrakech region (Morocco) and analysis of the otsA gene putatively involved in osmotolerance. Syst Appl Microbiol 35: 156–164. doi: 10.1016/j.syapm.2012.02.003
|
| [101] | Mnasri B, Mrabet M, Laguerre G (2007) Salt-tolerant rhizobia isolated from a Tunisian oasis that are highly effective for symbiotic N2-fixation with Phaseolus vulgaris constitute a novel biovar (bv. mediterranense) of Sinorhizobium meliloti. Arch Microbiol 187: 79–85. |
| [102] | Mnasri B, Saïdi S, Chihaouri SA, et al. (2012) Sinorhizobium americanum symbiovar mediterranense is a predominant symbiont that nodulates and fixes nitrogen with common bean (Phaseolus vulgaris L.) in a Northern Tunisian field. Syst Appl Microbiol 35: 263–269. |
| [103] | Shamseldin AAY, Vinuesa P, Thierfelder H, et al. (2005) Rhizobium etli and Rhizobium gallicum nodulate Phaseolus vulgaris in Egyptian soils and display cultivar-dependent symbiotic efficiency. Symbiosis 38: 145–161. |
| [104] |
Shamseldin A, Werner D (2005) High salt and high pH tolerance of new isolated Rhizobium etli strains from Egyptian soils. Curr Microbiol 50: 11–16. doi: 10.1007/s00284-004-4391-7
|
| [105] | Anyango B, Wilson KJ, Beynon JL, et al. (1995) Diversity of rhizobia nodulating Phaseolus vulgaris L. in two Kenyan soils with contrasting pHs. Appl Environ Microbiol 61: 4016–4021. |
| [106] | Kawaka F, Dida MM, Opala PA, et al. (2014) Symbiotic efficiency of native rhizobia nodulating common bean (Phaseolus vulgaris L.) in soils of Western Kenya. Int Sch Res Notices. |
| [107] | Waswa NM (2014) Identifying elite rhizobia for commercial soybean (Glycine max) inoculants [MS Thesis], University of Nairobi, Nairobi, Kenya. |
| [108] |
Zinga M, Jaiswat SK, Dakora FD (2017) Presence of diverse rhizobial communities responsible for nodulation of common bean (Phaseolus vulgaris) in South African and Mozambican soils. FEMS Microbiol Ecol 93: fiw236. doi: 10.1093/femsec/fiw236
|
| [109] | Gurkanli CT, Ozkoc I, Gunduz I (2013) Genetic diversity in rhizobia nodulating common bean (Phaseolus vulgaris L.) in the Central Black Sea Region of Turkey. Ann Microbiol 63: 971–987. |
| [110] |
Tamimi SM, Young JPW (2004) Rhizobium etli is the dominant common bean nodulating rhizobia in cultivated soils from different locations in Jordan. Appl Soil Ecol 26: 193–200. doi: 10.1016/j.apsoil.2004.01.003
|
| [111] |
Assadi Rahmani H, Rasanen LA, Afshari A, et al. (2011) Genetic diversity and symbiotic effectiveness of rhizobia isolated from root nodules of Phaseolus vulgaris grown in soils of Iran. Appl Soil Ecol 48: 287–293. doi: 10.1016/j.apsoil.2011.04.010
|
| [112] |
Abbaszadeh-dahaji P, Savaghebi RG, Asadi-Rahmani H, et al. (2012) Symbiotic effectiveness and plant growth promoting traits in some Rhizobium strains isolated from Phaseolus vulgaris L. Plant Growth Regul 68: 361–370. doi: 10.1007/s10725-012-9724-0
|
| [113] | Zhang X, Blair MW, Wang S (2008) Genetic diversity of Chinese common bean (Phaseolus vulgaris L.) landraces assessed with simple sequence repeat markers. Theor Appl Genet 117: 629–640. |
| [114] | Van Scoonhoven A, Voyseste O (1991) Common beans: research for crop improvement, CAB International Publishing. |
| [115] |
Han SZ, Wang ET, Chen WX (2005) Diverse bacteria isolated from root nodules of Phaseolus vulgaris and species withinthe genera Campylotropis and Cassia grown in China. Syst Appl Microbiol 28: 265–276. doi: 10.1016/j.syapm.2004.12.005
|
| [116] |
Wang L, Cao Y, Wang ET (2016) Biodiversity and biogeography of rhizobia associated with common bean (Phaseolus vulgaris L.) in Shaanxi Province. Syst Appl Microbiol 39: 211–219. doi: 10.1016/j.syapm.2016.02.001
|
| [117] |
Cao Y, Wang ET, Zhao L, et al. (2014) Diversity and distribution of rhizobia nodulated with Phaseolus vulgaris in two ecoregions of China. Soil Biol Biochem 78: 128–137. doi: 10.1016/j.soilbio.2014.07.026
|
| [118] | Adhikari D, Itoh K (2013) Genetic diversity of common bean (Phaseolus vulgaris L.) nodulating rhizobia in Nepal. Plant Soil 368: 341–353. |
| [119] | Herridge DF, Redden RJ (1999) Evaluation of genotypes of navy and culinary bean (Phaseolus vulgaris L.) selected for superior growth and nitrogen fixation. Aust J Exp Agric 39: 975–980. |
| [120] | Kellman AW, Hill GD, McKenzie BA (2006) Is it worth inoculating common bean (Phaseolus vulgaris L.)? Agronomy, NZ 36: 24–35. |
| [121] |
Lafay B, Burdon JJ (2007) Molecular diversity of legume root-nodule bacteria in Kakadu National Park, Northern Territory, Australia. PLoS One 2: e277. doi: 10.1371/journal.pone.0000277
|
| [122] | Eardly B, Elia P, Brockwell J, et al. (2017) Biogeography of a novel Ensifer meliloti clade associated with the Australian legume Trigonella suavissima. Appl Environ Microbiol 83: e03446-16. |
| [123] |
Young JPW, Johnston AWB (1989) The evolution of specificity in the legume-rhizobium symbiosis. Trends Ecol Evol 4: 341–349. doi: 10.1016/0169-5347(89)90089-X
|
| [124] |
Wang D, Yang S, Tang F (2012) Symbiosis specificity in the legume-rhizobial mutualism. Cell Microbiol 14: 334–342. doi: 10.1111/j.1462-5822.2011.01736.x
|
| [125] |
Dalla Via V, Zanetti ME, Blanco F (2016) How legumes recognize rhizobia. Plant Signal Behav 11: e1120396-5. doi: 10.1080/15592324.2015.1120396
|
| [126] |
Cooper JE (2007) Early interactions between legumes and rhizobia: disclosing complexity in a molecular dialogue. J Appl Microbiol 103: 1355–1365. doi: 10.1111/j.1365-2672.2007.03366.x
|
| [127] |
Janczarek M (2011) Environmental signals and regulatory pathways that influence exopolysaccharide production in hizobia. Int J Mol Sci 12: 7898–7933 doi: 10.3390/ijms12117898
|
| [128] | Sutherland IW (1972) Bacterial exopolysaccharides. AdvMicrob Physiol 8: 143–213. |
| [129] |
Gibson KE, Kobayashi H, Graham C, et al. (2008) Molecular determinants of a symbiotic chronic infection. Annu Rev Genet 42: 413–441. doi: 10.1146/annurev.genet.42.110807.091427
|
| [130] |
Long SR (1996) Rhizobium symbiosis: Nod factors in perspective. Plant Cell 8: 1885–1898. doi: 10.1105/tpc.8.10.1885
|
| [131] |
Mergaert P, Van Montagu M, Holsters M (1997) Molecular mechanisms of Nod factor diversity. Mol Microbiol 25: 811–817. doi: 10.1111/j.1365-2958.1997.mmi526.x
|
| [132] | Bolaños-Vásquez MC, Werner D (1997) Effect of Rhizobium tropici, R. etli, and R. leguminosarum bv. phaseoli on nod gene inducing flavonoids in root exudates of Phaseolus vulgaris. Mol Plant Microbe In 10: 339–346. |
| [133] |
Martínez-Romero E (2009) Coevolution in Rhizobium-legume symbiosis? DNA Cell Biol 28: 361–370. doi: 10.1089/dna.2009.0863
|
| [134] | Jorrín B, Imperial J (2015) Pool-seq analysis of microsymbiont selection by the legume plant host, In: de Brujin FJ, Biological Nitrogen Fixation, Hoboken: John Wiley & Sons, Inc. |
| [135] |
Jorrín B, Imperial J (2015) Population genomics analysis of legume host preference for specific rhizobial genotypes in the Rhizobium leguminosarum bv. viciae symbioses. Mol Plant Microbe In 28: 310–318. doi: 10.1094/MPMI-09-14-0296-FI
|
| [136] |
Reid DE, Ferguson BJ, Hayashi S, et al. (2011) Molecular mechanisms controlling legume autoregulation of nodulation. Ann Bot 108: 789–795. doi: 10.1093/aob/mcr205
|
| [137] |
Formey D, Martín-Rodríguez JÁ, Leija A, et al. (2016) Regulation of small RNAs and corresponding targets in Nod factor-induced Phaseolus vulgaris root hair cells. Int J Mol Sci 17: 887. doi: 10.3390/ijms17060887
|
| [138] |
Vargas C, Martínez LJ, Megías M, et al. (1990) Identification and cloning of nodulation genes and host specificity determinants of the broad host-range Rhizobium leguminosarum strain CIAT899. Mol Microbiol 4: 1899–1910. doi: 10.1111/j.1365-2958.1990.tb02039.x
|
| [139] |
Yang GP, Debellé F, Savagnac A, et al. (1999) Structure of the Mesorhizobium hualuii and Rhizobium galegae Nod factors: a cluster of phylogenetically related legumes are nodulated by rhizobia producing Nod factors with α, β-unsaturated N-acyl substitutions. Mol Microbiol 34: 227–237. doi: 10.1046/j.1365-2958.1999.01582.x
|
| [140] |
Soria-Díaz ME, Rodríguez-Carvajal MA, Tejero-Mateo P, et al. (2006) Structural determination of the Nod factors produced by Rhizobium gallicum bv. gallicum R602. FEMS Microbiol Lett 255: 164–173. doi: 10.1111/j.1574-6968.2005.00065.x
|
| [141] |
Del Cerro P, Rolla-Santos AA, Gomes DF, et al. (2015) Opening the "black box" of nodD3, nodD4 and nodD5 genes of Rhizobium tropici strain CIAT899. BMC Genomics 16: 864. doi: 10.1186/s12864-015-2033-z
|
| [142] |
Del Cerro P, Rolla-Santos AA, Gomes DF, et al. (2015) Regulatory nodD1 and nodD2 genes of Rhizobium tropici strain CIAT899 and their roles in the early stages of molecular signaling and host-legume nodulation. BMC Genomics 16: 251. doi: 10.1186/s12864-015-1458-8
|
| [143] |
Prudent M, Salon C, Smith DL (2016) Nod factor supply under water stress conditions modulates cytokinin biosynthesis and enchances nodule formation and N nutrition in soybean. Plant Signal Behav 11: e1212799. doi: 10.1080/15592324.2016.1212799
|
| [144] |
Lafuente A, Pérez-Palacios P, Doudkali B, et al. (2015) Unraveling the effect of arsenic on the model Medicago-Ensifer interaction: a transcriptomic meta-analysis. New Phytol 205: 255–272. doi: 10.1111/nph.13009
|
| [145] | Duzan HM, Zhou X, Souleimanov A, et al. (2004) Perception of Bradyrhizobium japonicum Nod factor in soybean [Glycine max (L.) Merr.] root hairs under abiotic stress conditions. J Exp Bot 55: 2641–2646. |
| [146] | de Souza EM, Granada CE, Sperotto RA (2016) Plant pathogens affecting the establishment of Plant-Symbiont Interaction. Front Plant Sci 7: 15. |
| [147] |
Perez-Carrascal OM, Vanlsberghe D, Juarez S, et al. (2016) Population genomics of the symbiotic plasmids of sympatric nitrogen-fixing Rhizobium species associated with Phaseolus vulgaris. Environ Microbiol 18: 2660–2676. doi: 10.1111/1462-2920.13415
|
| [148] |
Laguerre G, Nour SM, Macheret V, et al. (2001) Classification of rhizobia based on nodC and nifH gene analysis reveals a close phylogenetic relationship among Phaseolus vulgaris symbionts. Microbiology 147: 981–993. doi: 10.1099/00221287-147-4-981
|
| [149] |
Cervantes L, Bustos P, Girard L, et al. (2011) The conjugative plasmid of a bean-nodulating Sinorrhizobium fredii strain is assembled from sequences of two Rhizobium strains and the chromosome of a Sinorhizobium strain. BMC Microbiol 11: 149. doi: 10.1186/1471-2180-11-149
|
| [150] |
Resendis-Antonio O, Hernandez M, Salazae E, et al. (2011) System biology of bacterial nitrogen fixation: high throughput technology and its integrative description with constraint-based modeling. BMC Syst Biol 5: 120. doi: 10.1186/1752-0509-5-120
|
| [151] | Gomes DF, Batista JS, Schiavon AL, et al. (2012) Proteomic profiling of Rhizobium tropici PRF 81: identification of conserved and specific responses to heat stress. BMC Microbiol 12: 84. |
| [152] |
Limpens E, Franken C, Smit P, et al. (2003) LysM domain receptor kinases regulating rhizobial Nod factor-induced infection. Science 302: 630–633. doi: 10.1126/science.1090074
|
| [153] |
Sato S, Nakamura Y, Kaneko T, et al. (2008) Genome structure of the legume Lotus japonicas.DNA Res 15: 227–239. doi: 10.1093/dnares/dsn008
|
| [154] | Young ND, Debelle F, Oldroyd GE, et al. (2011) The Medicago genome provides insight into the evolution of rhizobial symbioses. Nature 480: 520–524. |
| [155] |
Schmutz J, Cannon SB, Schlueter J, et al. (2010) Genome sequence of the palaeopolyploid soybean. Nature 463: 178–183. doi: 10.1038/nature08670
|
| [156] |
Varshney RK, Song C, Saxena RK, et al. (2013) Draft genome sequence of chickpea (Cicer arietinum) provides a resource for trait improvement. Nat Biotechnol 31: 240–246. doi: 10.1038/nbt.2491
|
| [157] |
Schmutz J, McLean P, Mamidi S, et al. (2014) A reference genome for common bean and genome-wide analysis of dual domestications. Nat Genet 46: 707–713. doi: 10.1038/ng.3008
|
| [158] |
Vlasova A, Capella-Gutierrez S, Rendon-Anaya M, et al. (2016) Genome and transcriptome analysis of the Mesoamerican common bean and the role of gene duplications in establishing tissue and temporal specialization of genes. Genome Biol 17: 32. doi: 10.1186/s13059-016-0883-6
|
| [159] |
Lin JY, Stupar RM, Hans C, et al. (2010) Structural and functional divergence of a 1-Mb duplicated region in the soybean (Glycine max) genome and comparison to an orthologous region from Phaseolus vulgaris. Plant Cell 22: 2545–2561. doi: 10.1105/tpc.110.074229
|
| [160] |
McClean PE, Marnidi S, McConnell M, et al. (2010) Synteny mapping between common bean and soybean reveals extensive blocks of shared loci. BMC Genomics 11: 184–194. doi: 10.1186/1471-2164-11-184
|
| [161] |
Schlueter JA, Dixon P, Granger C, et al. (2004). Mining EST databases to resolve evolutionary events in major crop species. Genome 47: 868–876. doi: 10.1139/g04-047
|
| [162] | Kalavacharia V, Liu Z, Meyers BC, et al. (2011) Identification and analysis of common bean (Phaseolus vulgaris L.) transcriptomes by massively parallel pyrosequencing. BMC Plant Biol 11: 135. |
| [163] |
Endre G, Kereszt A, Kevei Z, et al. (2002) A receptor kinase gene regulating symbiotic nodule development. Nature 417: 962–966. doi: 10.1038/nature00842
|
| [164] |
Zhu H, Choi HK, Cook DR, et al. (2005). Bridging model and crop legumes through comparative genomics. Plant Physiol 137: 1189–1196. doi: 10.1104/pp.104.058891
|
| [165] | Kim DH, Parupalli S, Azam S, et al. (2013) Comparative sequence analysis of nitrogen fixation-related genes in six legumes. Front Plant Sci 4: 300. |
| [166] | Qiao Z, Pingault L, Nourbakhsh-Rey M, et al. (2016) Comprehensive comparative genomic and transcriptomic analyses of the legume genes controlling the nodulation process. Front Plant Sci 7: 34. |
| [167] |
Ferguson BJ, Indrasumunar A, Hayashi S, et al. (2010) Molecular analysis of legume nodule development and autorregulation. J Integr Plant Biol 52: 61–76. doi: 10.1111/j.1744-7909.2010.00899.x
|
| [168] | Shritliffe SJ, Vessey JK, Buttery, et al. (1996) Comparison of growth and N accumulation of common bean (Phaseolus vulgaris L.), cv. OAC Rico and its two nodulation mutants, R69 and R99. Can J Plant Sci 76: 73–83. |
| [169] | Abd-Alla MH (2011) Nodulation and nitrogen fixation in interspecies grafts of soybean and common bean is controlled by isoflavonoid signal molecules translocated from shoot. Plant Soil Environ 57: 453–458. |
| [170] | Hastwell AH, Gresshoff PM, Ferguson BJ (2015) Genome-wide annotation and characterization of CLAVATA/ESR (CLE) peptide hormones of soybean (Glycine max) and common bean (Phaseolus vulgaris), and their orthologues of Arabidopsis thaliana. JExp Bot 66: 5271–5287. |
| [171] |
Ferguson BJ, Mathesius U (2014) Phytohormone regulation of legume-rhizobia interactions. J Chem Ecol 40: 770–790. doi: 10.1007/s10886-014-0472-7
|
| [172] |
Oldroyd GED, Downie JA (2008) Coordinating nodule morphogenesis with rhizobial infection in legumes. Annu Rev Plant Biol 59: 519–546. doi: 10.1146/annurev.arplant.59.032607.092839
|
| [173] |
Reid DE, Ferguson BJ, Gresshoff PM (2011) Inoculation- and nitrate-induced CLE peptides of soybean control NARK-dependent nodule formation. Mol Plant Microbe In 24: 606–618. doi: 10.1094/MPMI-09-10-0207
|
| [174] |
O'Rourke JA, Iniguez LP, Fu F, et al. (2014) An RNA-Seq based gene expression atlas of the common bean. BMC Genomics 15: 866. doi: 10.1186/1471-2164-15-866
|
| [175] | Carroll BJ, McNeil DL, Gresshoff PM (1985) Isolation and properties of soybean [Glycine max (L.) Merr.] mutants that nodulate in the presence of high nitrate concentrations. Proc Natl Acad Sci USA 82: 4162–4166. |
| [176] |
Carroll BJ, McNeil DL, Gresshoff PM (1985) A supernodulation and nitrate tolerant symbiotic (nts) soybean mutant. Plant Physiol 78: 34–40. doi: 10.1104/pp.78.1.34
|
| [177] | Gresshoff PM (1993) Molecular genetic analysis of nodulation genes in soybean. Plant Breed Rev 11: 275–318. |
| [178] |
Szczyglowski K, Shaw RS, Wopereis J, et al. (1998) Nodule organogenesis and symbiotic mutants of the model legume Lotus japonicus. Mol Plant Microbe In 11: 684–697. doi: 10.1094/MPMI.1998.11.7.684
|
| [179] |
Zanetti ME, Blanco FA, Beker MP, et al. (2010) A C subunit of the plant nuclear factor NF-Y required for rhizobial infection and nodule development affects partner selection in the common bean-Rhizobium etli symbiosis. Plant Cell 22: 4142–4157. doi: 10.1105/tpc.110.079137
|
| [180] |
Ramírez M, Graham MA, Blanco-López L (2005) Sequencing and analysis of common bean ESTs: Building a foundation for functional genomics. Plant Physiol 137:1211–1227. doi: 10.1104/pp.104.054999
|
| [181] |
Peltzer Meschini EP, Blanco FA, Zanetti ME, et al. (2008). Host genes involved in nodulation preference in common bean (Phaseolus vulgaris)-Rhizobium etli symbiosis revealed by suppressive subtractive hybridization. Mol Plant Microbe In 21: 459–468. doi: 10.1094/MPMI-21-4-0459
|
| [182] | Galeano CH, Cortés AJ, Fernandez AC (2012) Gene-based single nucleotide polymorphism markers for genetic and association mapping in common bean. BMC Genet 13: 48. |
| [183] | Quiceno-Rico JM, Camas-Reyes JL, Alvarez-Venegas R (2012) Molecular cloning and characterization of two trithorax-group genes from Phaseolus vulgaris roots and symbiotic nodules. Plant Omics 5: 103–114. |
| [184] | Montiel J, Arthikala MK, Quinto C (2013) Phaseolus vulgarisRbohB functions in lateral root development. Plant Signal Behav 1: 8. |
| [185] | Islas-Flores T, Guillén G, Alvarado-Affantranger X, et al. (2011) PvRACK1 loss-of-function impairs expansion and morphogenesis in Phaseolus vulgaris L. root nodules. Mol Plant Microbe In 24: 819–826. |
| [186] | Barraza A, Estrada-Navarrete G, Rodriguez-Algeria ME, et al. (2013) Down-regulation of PvTRE1 enhances nodule biomass and bacteriod number in the common bean. New Phytol 197: 194–206. |
| [187] |
Dalla Via V, Narduzzi C, Aguilar OM, et al. (2015) Changes in the common bean transcriptome in response to secreted and surface signal molecules of Rhizobium etli. Plant Physiol 169: 1356–1370. doi: 10.1104/pp.15.00508
|
| [188] | Yan Z, Hossain MS, Wang J, et al. (2013) miR172 regulates soybean nodulation. Mol Plant Microbe In 26: 1371–1377. |
| [189] |
Wang Y, Wang L, Zou Y (2014) Soybean miR172c targets the repressive AP2 transcription factor NNC1 to activate ENOD40 expression and regulate nodule initiation. Plant Cell 26: 4782–4801. doi: 10.1105/tpc.114.131607
|
| [190] | Nova-Franco B, Íñiguez LP, Valdés-López O (2015) The miR172c-AP2-1 node as a key regulator of the common bean-rhizobia nitrogen fixation symbiosis. Plant Physiol 68: 273–291. |
| [191] |
Buttery BR, Park SJ, Berkum PV (1997) Effects of common bean (Phaseolus vulgaris L) cultivar and Rhizobum strain on plant growth, seed yield and nitrogen content. Can J Plant Sci 77: 347–351. doi: 10.4141/P96-088
|
| [192] |
Elizondo Barron J, Pasino RJ, Davis DW, et al. (1999) Response to selection for seed yield and nitrogen (N2) fixation in common bean (Phaseolus vulgaris L.). Field Crops Res 62: 119–128. doi: 10.1016/S0378-4290(99)00009-X
|
| [193] | Graham PH, Rosas J (1977) Growth and development of indeterminate bush and climbing cultiuvars of Phaseolus vulgaris L. inoculated with Rhizobium.J Agric Sci 88: 503–508. |
| [194] |
Pereira PAA, Miranda BD, Attewell JR, et al. (1993) Selection for increased nodule number in common bean (Phaseolus vulgaris L.). Plant Soil 148: 203–209. doi: 10.1007/BF00012858
|
| [195] | Rennie RJ, Kemp GA (1983) N2-fixation in field beans quantified by 15N isotope dilution. II. Effect of cultivars of beans. Agron J 75: 645–649. |
| [196] | Buttery BR, Park SJ, van Berkum P (1997) Effects of common bean (Phaseolus vulgaris L.) cultivar and rhizobium strain on plant growth, seed yield and nitrogen content. CanJ Plant Sci 77: 347–351. |
| [197] |
Rodiño AP, De La Fuente M, De Ron AM, et al. (2011) Variation for nodulation and plant yield of common bean genotypes and environmental effects on the genotype expression. Plant Soil 346: 349–361. doi: 10.1007/s11104-011-0823-x
|
| [198] | Rodiño AP, Santalla M, De Ron AM, et al. (2005) variability in symbiotic nitrogen fixation among white landraces of common bean from the Iberian peninsula. Symbiosis 40: 69–78. |
| [199] |
Polania J, Poschenrieder C, Rao I, et al. (2016) Estimation of phenotypic variability in symbiotic nitrogen fixation ability in common bean under drought stress using 15N natural abundance in grain. Eur J Agron 79: 66–73. doi: 10.1016/j.eja.2016.05.014
|
| [200] | Nodari RO, Tsai SM, Guzman P, et al. (1993) Towards an integrated linkage map of common bean III: Mapping genetic factors controlling host-bacteria interactions. Genetics 134: 341–350. |
| [201] | Tsai S, Nodari R, Moon D, et al. (1998) QTL mapping for nodule number and common bacterial blight in Phaseolus vulgaris L. Plant Soil 204: 135–145. |
| [202] |
Souza A, Boscariol R, Moon D, et al. (2000) Effects of Phaseolus vulgaris QTL in controlling host-bacteria interactions under two levels of nitrogen fertilization. Gen Mol Biol 23: 155–161. doi: 10.1590/S1415-47572000000100029
|
| [203] |
Ramaekers L, Galeano C, Garzón N, et al. (2013) Identifying quantittative trait loci for symbiotic nitrogen fixation capacity and related traits in common bean. Mol Breeding 31: 163–180. doi: 10.1007/s11032-012-9780-1
|
| [204] | Kamfwa K, Cichy KA, Kelly JD (2015) Genome-wide association analysis of symbiotic nitrogen fixation in common bean. Theor Appl Genet 128: 2017. |
| [205] |
Dwivedi SL, Sahrawat KL, Upadhyaya HD, et al. (2015) Advances in host plant and rhizobium genomics to enhance symbiotic nitrogen fixation in grain legumes. Adv Agron 129: 1–116. doi: 10.1016/bs.agron.2014.09.001
|
| [206] | Franco MC, Cassini ST, Oliveira VR, et al. (2001) Combining ability for nodulation in common bean (Phaseolus vulgaris L.) genotypes from Andean and middle American gene pools. Euphytica 118: 265–270. |
| [207] | Asfaw A, Blair MW, Struik P (2012) Multi-environment quantitative trait loci analysis for photosynthate acquisition, accumulation, and remobilization traits in common bean under drought stress. G3 Genes Genom Genet 2: 579–595. |
| [208] | Farid M (2015) Symbiotic nitrogen fixation in common bean, PhD Thesis Dissertation, University of Guelph, Ontario. |
| [209] |
Heilig JA, Beaver JS, Wright EM, et al. (2017) QTL analysis of symbiotic nitrogen fixation in a black bean population. Crop Sci 57: 118–129. doi: 10.2135/cropsci2016.05.0348
|
| [210] |
Sánchez-López, Jáuregui D, Nava N (2011) Down-regulation of SymRK correlates with a deficiency in vascular bundle development in Phaseolus vulgaris nodules. Plant Cell Environ 34: 2109–2121. doi: 10.1111/j.1365-3040.2011.02408.x
|
| [211] |
Mitra RM, Gleason CA, Edwards A, et al. (2004) A Ca2+/calmodulin-dependent protein kinase required for symbiotic nodule development: gene identification by transcript-based cloning. Proc Natl Acad Sci USA 101: 4701–4705. doi: 10.1073/pnas.0400595101
|
| [212] | Pedalino MJ, Kipe-Nolt L (1993) Common bean (Phaseolus vulgaris L.) mutants defective in root nodule formation. I. Physiological characterization. J Exp Bot 44: 1007–1014. |
| [213] | Pedalino MJ, Kipe-Nolt L, Frusciante L, et al. (1993) Common bean (Phaseolus vulgaris L.) mutants defective in root nodule formation. II. Genetic analysis. J Exp Bot 44: 1015–1020. |
| [214] | Park SJ, Buttery BR (1988) Nodulation mutants of white bean (Phaseolus vulgaris L.), induced by ethyl-methane sulphonate. Can J Plant Sci 68: 199–202. |
| [215] |
Park SJ, Buttery BR (1997) Complementation of nodulation genes of various mutants in common bean (Phaseolus vulgaris L.). J Hered 88: 543–545. doi: 10.1093/oxfordjournals.jhered.a023155
|
| [216] |
Pedalino M, Giller KE, Kipe-Nolt J (1992) Genetics of physiological characterization of non-nodulating mutant of Phaseolus vulgaris L.-NOD125. J Exp Bot 43: 843–849. doi: 10.1093/jxb/43.6.843
|
| [217] | Park SJ, Buttery BR (1989) Identification and characterization of common bean (Phaseolus vulgaris L.) lines well nodulated in the presence of high nitrate. Plant Soil 119: 237–244. |
| [218] | Park SJ, Buttery BR (1994) Inheritance of non-nodulation and ineffective nodulation mutants in common bean. J Hered 85: 1–3. |
| [219] | Shritliffe SJ, Vessey JK (1996) A nodulation (Nod+/Fix-) mutant of Phaseolus vulgaris L. has nodule-like structures lacking peripheral vascular bundles (Pvb-) and is resistant to mycorrhizal infection (Myc-). Plant Sci 118: 209–220. |
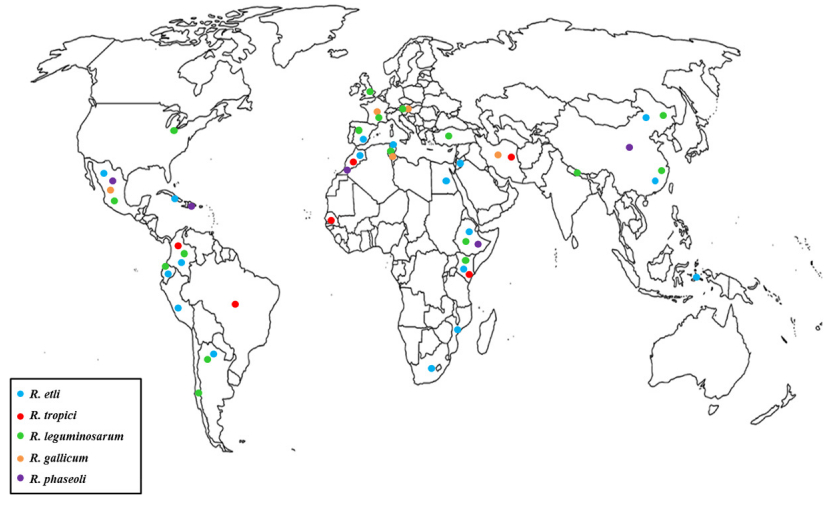
Figures(1) / Tables(2)

Olaya Muñoz-Azcarate, Ana M González, Marta Santalla. Natural rhizobial diversity helps to reveal genes and QTLs associated with biological nitrogen fixation in common bean[J]. AIMS Microbiology, 2017, 3(3): 435-466. doi: 10.3934/microbiol.2017.3.435







 DownLoad:
DownLoad: