Citation: Vanessa J. White, Ramesh C. Nayak. Re-circulating Phagocytes Loaded with CNS Debris: A Potential Marker of Neurodegeneration in Parkinsons Disease?[J]. AIMS Medical Science, 2015, 2(1): 26-34. doi: 10.3934/medsci.2015.1.26

| [1] |
Berg D, Marek K, Ross GW, Poewe W (2012). Defining At-Risk Populations for Parkinson's disease: Lessons from Ongoing Studies. Movement Disord 27: 656-665. doi: 10.1002/mds.24985

|
| [2] | Spiegel J, Storch A, Jost WH (2006). Early diagnosis of Parkinson's disease. J Neurol 253 Suppl4(S4): IV2-7. |
| [3] |
Shi C, Pamer EG (2011). Monocyte recruitment during infection and inflammation. Nat Rev Immunol 11: 762-774. doi: 10.1038/nri3070
|
| [4] |
van der Goes A, Boorsmaa W, Hoekstraa K, et al. (2005). Determination of the sequential degradation of myelin proteins by macrophages. J Neuroimmunol 161: 12-20. doi: 10.1016/j.jneuroim.2004.12.010
|
| [5] |
Hoehn M, Yahr M (1967). Parkinsonism: onset, progression and mortality. Neurology 17: 427-42. doi: 10.1212/WNL.17.5.427
|
| [6] |
Rosas AL, Nosanchuk JD, Casadevall (2001) Passive Immunization with Melanin-Binding Monoclonal Antibodies Prolongs Survival of Mice with Lethal Cryptococcus neoformans Infection. Infect Immun 69: 3410-3412. doi: 10.1128/IAI.69.5.3410-3412.2001
|
| [7] |
Dadachova E, Moadel T, Schweitzer AD, et al. (2006) Radiolabeled Melanin-Binding Peptides Are Safe and Effective in Treatment of Human Pigmented Melanoma in a Mouse Model of Disease. Cancer Biother Radio 21: 117-129. doi: 10.1089/cbr.2006.21.117
|
| [8] |
Lee V M-Y, Goedert M, Trojanowski JQ (2001) Neurodegenerative Tauopathies. Annu Rev Neurosci 24: 1121-1159. doi: 10.1146/annurev.neuro.24.1.1121
|
| [9] | Burgoyne RD (2007). Neuronal Calcium Sensor Proteins: Generating Diversity in Neuronal Ca2+Signalling. Nat Rev Neurosci 8: 182-193 |
| [10] | Doran JF, Jackson P, Kynoch PA, et al. (1983). Isolation of PGP 9. 5, a new human neurone-specific protein detected by high-resolution two-dimensional electrophoresis. J Neurochem 40: 1542-1547 |
| [11] |
Maraganore DM, Lesnick TG, Elbaz A, et al. (2004). UCHL1 is a Parkinson's disease susceptibility gene. Ann Neurol 55: 512-21. doi: 10.1002/ana.20017
|
| [12] |
Leroy E, Boyer R, Auburger G, et al. (1998). The ubiquitin pathway in Parkinson's disease. Nature 395: 451-2. doi: 10.1038/26652
|
| [13] |
Harhangi BS, Farrer MJ, Lincoln S, etal. (1999). The Ile93Met mutation in the ubiquitin carboxy-terminal-hydrolase-L1 gene is not observed in European cases with familial Parkinson's disease. Neurosci Lett 270: 1-4. doi: 10.1016/S0304-3940(99)00465-6
|
| [14] |
Joly S, Francke M, Ulbricht E, Beck S, et al. (2009) Cooperative Phagocytes: Resident Microglia and Bone Marrow Immigrants Remove Dead Photoreceptors in Retinal Lesions. Am J Pathol 174:2310-2323. doi: 10.2353/ajpath.2009.090023
|
| [15] | Peng T, Li C, Li X, et al. (2014). Correlation between Serum RANTES Levels and the Severity of Parkinson's Disease. Oxid Med Cell Longev 2014: 1-4. |
| [16] | Scalzo P, Silva de Miranda A, Guerra Amaral DC et al. (2011). Serum levels of chemokines in Parkinson's disease. Neuroimmunomodulat 23: 240-4 |
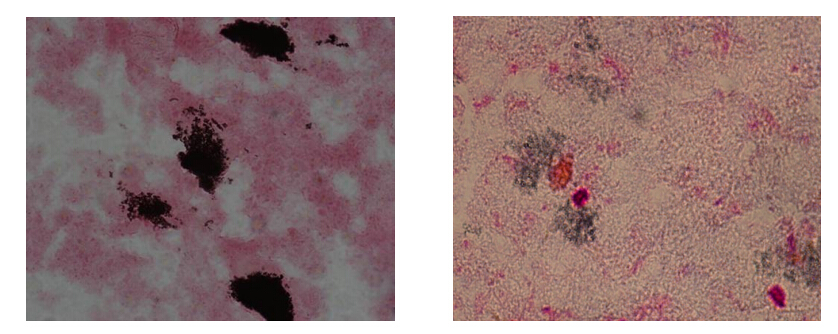
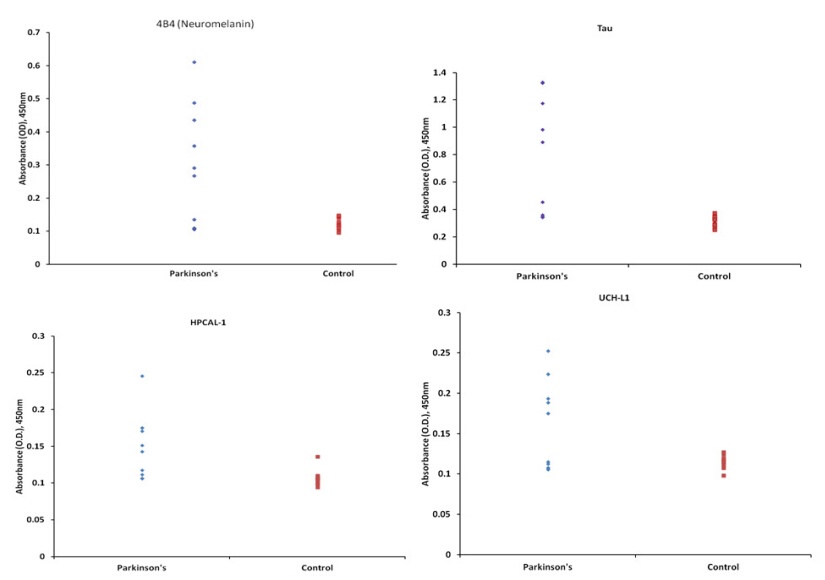
Figures(2)

Vanessa J. White, Ramesh C. Nayak. Re-circulating Phagocytes Loaded with CNS Debris: A Potential Marker of Neurodegeneration in Parkinsons Disease?[J]. AIMS Medical Science, 2015, 2(1): 26-34. doi: 10.3934/medsci.2015.1.26







 DownLoad:
DownLoad: