Citation: Nicola Vassena. Good and bad children in metabolic networks[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7621-7644. doi: 10.3934/mbe.2020388

| [1] | F. Horn, R. Jackson, General mass action kinetics, Arch. Ration. Mech. Anal., 47 (1972), 81-116. |
| [2] | M. Feinberg, Chemical reaction network structure and the stability of complex isothermal reactors—I. The deficiency zero and deficiency one theorems, Chem. Eng. Sci., 42 (1987), 2229-2268. |
| [3] |
M. Feinberg, The existence and uniqueness of steady states for a class of chemical reaction networks, Arch. Ration. Mech. Anal., 132 (1995), 311-370. doi: 10.1007/BF00375614

|
| [4] |
D. Gale, H. Nikaido, The Jacobian matrix and global univalence of mappings, Math. Ann., 159 (1965), 81-93. doi: 10.1007/BF01360282
|
| [5] |
G. Craciun, M. Feinberg, Multiple equilibria in complex chemical reaction networks: II. The species-reaction graph, SIAM J. Appl. Math., 66 (2006), 1321-1338. doi: 10.1137/050634177
|
| [6] | M. Banaji, P. Donnell, S. Baigent, P matrix properties, injectivity, and stability in chemical reaction systems, SIAM J. Appl. Math., 67 (2007), 1523-1547. |
| [7] |
M. Banaji, G. Craciun, Graph-theoretic criteria for injectivity and unique equilibria in general chemical reaction systems, Adv. Appl. Math., 44 (2010), 168-184. doi: 10.1016/j.aam.2009.07.003
|
| [8] |
G. Shinar, M. Feinberg, Concordant chemical reaction networks, Math. Biosci., 240 (2012), 92-113. doi: 10.1016/j.mbs.2012.05.004
|
| [9] |
G. Shinar, M. Feinberg, Concordant chemical reaction networks and the species-reaction graph, Math. Biosci., 241 (2013), 1-23. doi: 10.1016/j.mbs.2012.08.002
|
| [10] | R. Thomas, On the relation between the logical structure of systems and their ability to generate multiple steady states or sustained oscillations, Numerical methods in the study of critical phenomena, Springer Series in Synergetics, vol 9. (1981) 180-193. |
| [11] |
C. Soulé, Graphic requirements for multistationarity, ComPlexUs, 1 (2003), 123-133. doi: 10.1159/000076100
|
| [12] |
M. Kaufman, C. Soulé, R. Thomas, A new necessary condition on interaction graphs for multi-stationarity, J. Theor. Biol., 248 (2007), 675-685. doi: 10.1016/j.jtbi.2007.06.016
|
| [13] |
C. Wiuf, E. Feliu, Power-law kinetics and determinant criteria for the preclusion of multistation-arity in networks of interacting species, SIAM J. Appl. Dyn. Syst., 12 (2013), 1685-1721. doi: 10.1137/120873388
|
| [14] | E. Feliu, N. Kaihnsa, T. de Wolff, O. Yürük, The kinetic space of multistationarity in dual phosphorylation, J. Dyn. Differ. Equ., (2020). |
| [15] | M. Mincheva, M. R. Roussel, Graph-theoretic methods for the analysis of chemical and biochemical networks. I. Multistability and oscillations in ordinary differential equation models, J. Math. Biol., 55 (2007), 61-86. |
| [16] | A. Ivanova, B. Tarnopolskii, One approach to the determination of a number of qualitative features in the behavior of kinetic systems, and realization of this approach in a computer (critical conditions, autooscillations), Kinet. Catal., 20 (1979), 1271-1277. |
| [17] | A. Volpert, A. Ivanova, Mathematical models in chemical kinetics, Math. Model. (Russian), 57 (1987), 102. |
| [18] | A. Ivanova, Conditions for uniqueness of the stationary states of kinetic systems, connected with the structure of their reaction-mechanism. 1., Kinet. Catal., 20 (1979), 833-837. |
| [19] |
B. Brehm, B. Fiedler, Sensitivity of chemical reaction networks: a structural approach. 3. Regular multimolecular systems, Math. Methods Appl. Sci., 41 (2018), 1344-1376. doi: 10.1002/mma.4668
|
| [20] | M. Feinberg, Foundations of Chemical Reaction Network Theory, Springer, 2019. |
| [21] | B. Fiedler, Global Hopf bifurcation in networks with fast feedback cycles, Discrete Contin. Dyn. Syst. Ser S, 0. |
| [22] | N. Vassena, Sensitivity of Metabolic Networks, PhD thesis, Freie Universität Berlin, 2020. |
| [23] | B. Alberts, D. Bray, J. Lewis, M. Raff, K. Roberts, J. D. Watson, Molecular biology of the cell. W.W. Norton & Co, 1983. |
| [24] | H. Lodish, A. Berk, C. A. Kaiser, M. Krieger, M. P. Scott, A. Bretscher, et al., Molecular cell biology, Macmillan, 2008. |
| [25] |
N. Ishii, K. Nakahigashi, T. Baba, M. Robert, T. Soga, A. Kanai, et al., Multiple high-throughput analyses monitor the response of e. coli to perturbations, Science, 316 (2007), 593-597. doi: 10.1126/science.1132067
|
| [26] |
K. Nakahigashi, Y. Toya, N. Ishii, T. Soga, M. Hasegawa, H. Watanabe, et al., Systematic phenome analysis of escherichia coli multiple-knockout mutants reveals hidden reactions in central carbon metabolism, Mol. Syst. Biol., 5 (2009), 306. doi: 10.1038/msb.2009.65
|
| [27] | F. G. Vital-Lopez, C. D. Maranas, A. Armaou, Bifurcation analysis of the metabolism of E. coli at optimal enzyme levels, in 2006 American Control Conference, IEEE, 2006, 6-pp. |
| [28] |
C. Chassagnole, N. Noisommit-Rizzi, J. W. Schmid, K. Mauch, M. Reuss, Dynamic modeling of the central carbon metabolism of Escherichia coli, Biotechnol. Bioeng., 79 (2002), 53-73. doi: 10.1002/bit.10288
|
| [29] |
T. Okada, J. C. Tsai, D. A. Mochizuki, Structural bifurcation analysis in chemical reaction networks, Phys. Rev. E, 98 (2018), 012417. doi: 10.1103/PhysRevE.98.012417
|
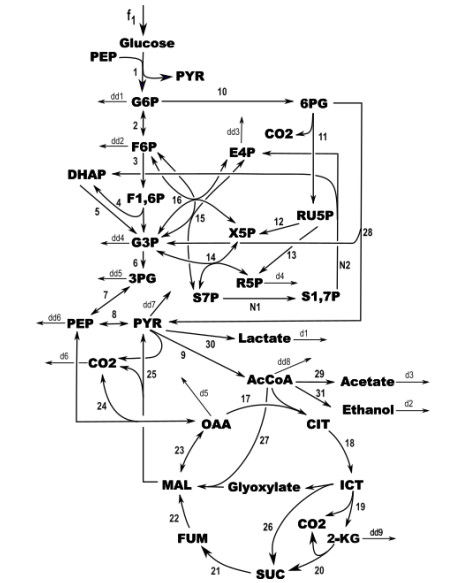
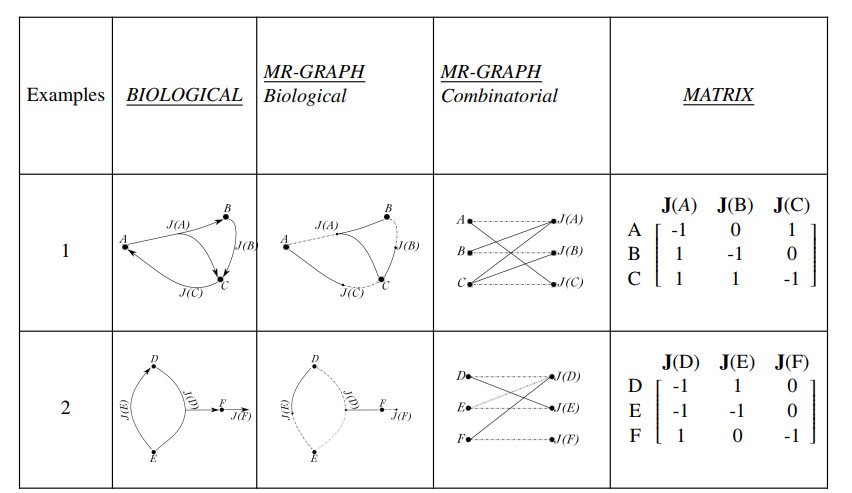
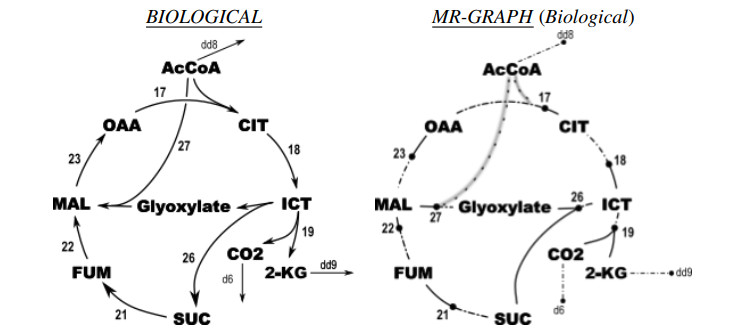
Figures(3)

Nicola Vassena. Good and bad children in metabolic networks[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7621-7644. doi: 10.3934/mbe.2020388







 DownLoad:
DownLoad: