Citation: Pengyi Hao, Sharon Chokuwa, Xuhang Xie, Fuli Wu, Jian Wu, Cong Bai. Skeletal bone age assessments for young children based on regression convolutional neural networks[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6454-6466. doi: 10.3934/mbe.2019323

| [1] | R. M. Malina and G. P. Beunen, Assessment of skeletal maturity and prediction of adult height (TW3 method), Am. J. Human Biol., 14(2002), 788–789. |
| [2] | D. R. Kirks and N. T. Griscom, Practical pediatric imaging: Diagnostic radiology of infants and children, Lippincott-Raven, (1998). |
| [3] | W. W. Greulich and S. I. Pyle, Radiographic atlas of skeletal development of the hand and wrist, Stanford University Press, (1959). |
| [4] | C. Athanasios, B. Maria, K. George, et al., Bone age estimation and prediction of final height in patients with β-thalassaemia major: a comparison between the two most common methods, Pediat. Radiol., 37(2007), 1241–1246. |
| [5] | T. S. Levitt, M. W. Hedgcock, J. W. Dye, et al., Bayesian inference for model-based segmentation of computed radiographs of the hand, Artif. Intell. Med., 5(1993), 365–387. |
| [6] | J. L. Zhang, P. Liu, F. Zhang, et al., CloudNet: Groundbased cloud classification with deep convolutional neural network, Geophys. Res. Lett., (2018), 8665–8672. |
| [7] | D. W. Zhang, D. Y. Meng and J. W. Han, Co-saliency detection via a self-paced multiple-instance learning framework, TPAMI, 39(2017), 865–878. |
| [8] | C. Bai, L. Huang, X. Pan, et al., Optimization of deep convolutional neural network for large scale image retrieval, Neurocomputing, 303(2018), 60–67. |
| [9] | X. He, H. Zhang, M. Landis, et al., Unsupervised boundary delineation of spinal neural foramina using a multi-feature and adaptive spectral segmentation, Med. Image Anal., 36(2017), 22–40. |
| [10] | C. Xu, L. Xu, Z. Gao, et al., Direct delineation of myocardial infarction without contrast agents using a joint motion feature learning architecture, Med. Image Anal., 50(2018), 82–94. |
| [11] | T. Xiao, L. Liu, K. Li, et al., Comparison of transferred deep neural networks in ultrasonic breast masses discrimination, BioMed. Res. Int., 36(2018), 1–9. |
| [12] | L. Hyunkwang, T. Shahein, S. Giordano, et al., Fully automated deep learning system for bone age assessment, J. Digit. Imaging, 30(2017), 427–441. |
| [13] | C. Spampinato, S. Palazzo, D. Giordano, et al., Deep learning for automated skeletal bone age assessment in X-Ray images, Med. Image Anal., 36(2017), 41–51. |
| [14] | X. Ren, T. Li, X. Yang, et al., Regression convolutional neural network for automated pediatric bone age assessment from hand radiograph, IEEE J. Biomed. Health, (2018). |
| [15] | S. Mutasa, P. D. Chang, C. Ruzal-Shapiro, et al., MABAL: a Novel Deep-Learning Architecture for Machine-Assisted Bone Age Labeling, J. Digit. Imaging, 31(2018), 513–519. |
| [16] | E. Pietka, L. Kaabi, M. L. Kuo, et al., Feature extraction in carpal-bone analysis, IEEE Trans. Med. Imaging, 12(1993), 44–49. |
| [17] | J. Seok, B. Hyun, J. Kasa-Vubu, et al., Automated classification system for bone age x-ray images, IEEE SMC, (2012), 208–213. |
| [18] | P. Liskowski and K. Krawiec, Segmenting retinal blood vessels with deep neural networks, IEEE Trans. Med. Imaging, 35(2016), 2369–2380. |
| [19] | K. Somkantha, N. Theera-Umpon and S. Auephanwiriyakul, Bone age assessment in young children using automatic carpal bone feature extraction and support vector regression, J. Digit. Imaging, 24(2011), 1044–1058. |
| [20] | C. W. Hsieh, A fuzzy-based growth model with principle component analysis selection for carpal bone-age assessment, Med. Biol. Eng. Comput., 48(2010), 579–588. |
| [21] | A. Zhang, A. Gertych and B. J. Liu, Automatic bone age assessment for young children from newborn to 7-year-old using carpal bones, Comput. Med. Imag. Grap., 31(2007), 299–310. |
| [22] | D. Giordano, C. Spampinato and G. Scarciofalo, An automatic system for skeletal bone age measurement by robust processing of carpal and epiphysial/metaphysial Bones, IEEE T. Instrum. Meas., 59(2010), 2539–2553. |
| [23] | P. Y. Hao, Y. J. Chen, S. Chokuwa, et al., Skeletal bone age assessment based on deep convolu-tional neural networks, PCM, (2018), 408–417. |
| [24] | S. Yadav and S. Shukla, Analysis of k-Fold cross-validation over hold-out validation on colossal datasets for quality classification, IACC, (2016), 78–83. |
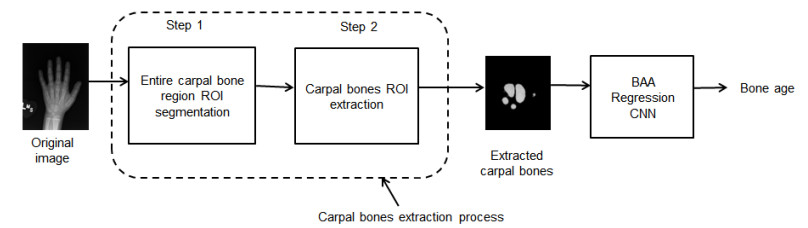
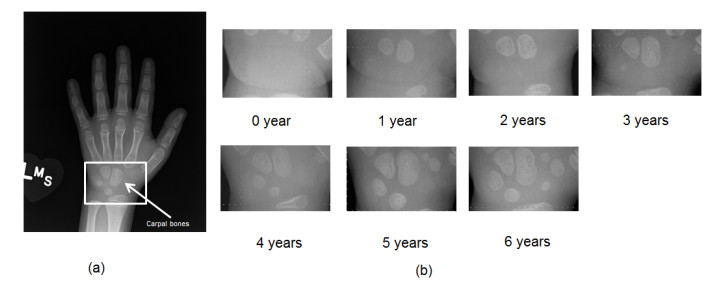
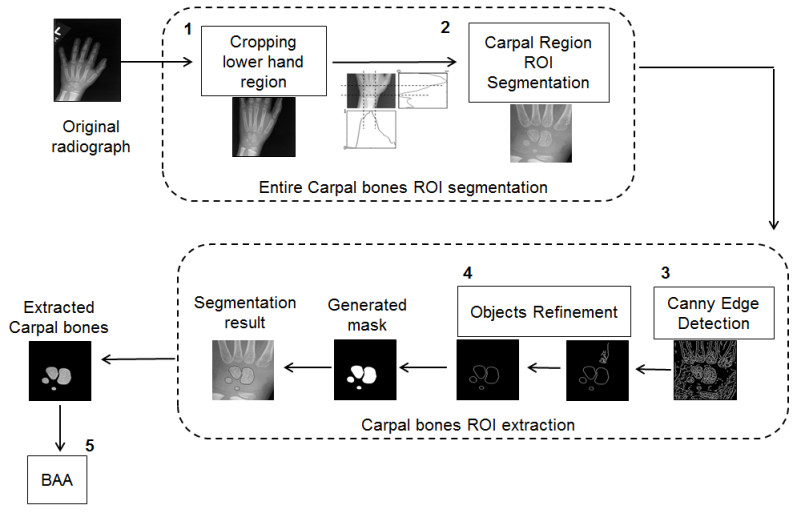
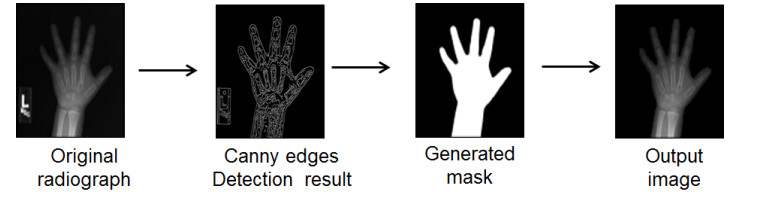
Figures(7)

Pengyi Hao, Sharon Chokuwa, Xuhang Xie, Fuli Wu, Jian Wu, Cong Bai. Skeletal bone age assessments for young children based on regression convolutional neural networks[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6454-6466. doi: 10.3934/mbe.2019323







 DownLoad:
DownLoad: