Citation: Xinyi Wang, He Wang, Shaozhang Niu, Jiwei Zhang. Detection and localization of image forgeries using improved mask regional convolutional neural network[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4581-4593. doi: 10.3934/mbe.2019229

| [1] | W. Luo, Z. Qu, F. Pan, et al., A survey of passive technology for digital image forensics, FCS, 1 (2007), 166–179. |
| [2] | J. G. R. Elwin, T. Aditya and S. M. Shankar, Survey on passive methods of image tampering detection, INCOCC,Erode, 2(2010), 431–436. |
| [3] | G. K. Birajdar and V. H. Mankar, Digital image forgery detection using passive techniques: A survey, Digit. Inve, 10 (2013), 226–245. |
| [4] | L. Verdoliva, D. Cozzolino and G. Poggi, A feature-based approach for image tampering detection and localization, WIFS, Atlanta, GA, (2014), 149–154. |
| [5] | U. H. Panchal and R. Srivastava, A comprehensive survey on digital image watermarking techniques, ICCSNT, (2015), 591–595. |
| [6] | Al-Qershi, M. Osamah and B. E. Khoo, Passive detection of copy-move forgery in digital images: State-of-the-art, Foren. Sci. Int., 23 (2013), 284–295. |
| [7] | H. Huang, W. Guo and Y. Zhang, Detection of copy-move forgery in digital images using SIFT algorithm, CIIA, (2008), 272–276. |
| [8] | T. Mahmood, A. Irtaza, Z. Mehmood, et al., Copy-move forgery detection through stationary wavelets and local binary pattern variance for forensic analysis in digital images, Forensic Sci. Int., 279 (2017), 8–21. |
| [9] | Y. Q. Shi, C. Chen and C. Wen, A natural image model approach to splicing detection, In Proceeding MM&Sec '07 Proceedings of the 9th workshop on Multimedia & security, (2007), 51–62. |
| [10] | X. Zhao, S. Wang, S, Li, et al., Passive image-splicing detection by a 2-D noncausal markov model, IEEE Trans. CSVT, 25 (2015), 185–199. |
| [11] | R. Girshick, Fast R-CNN, ICCV, (2015), 1440–1448. |
| [12] | B. H. Jawadul and A. K. Roy-Chowdhury, CNN based region proposals for efficient object detection, ICIP, (2016), 3658–3662. |
| [13] | B. Zhou, A. Lapedriza, J. Xiao, et al., Learning deep features for scene recognition using places database, ANIPS, 1(2015), 13–20 |
| [14] | L. Jonathan, E. Shelhamer and T. Darrell, Fully convolutional networks for semantic segmentation, IEEE Trans. Pattern Anal. Mach. Intel., 39 (2014), 640–651. |
| [15] | J. Chen, X. Kang, Y. Liu, et al., Median filtering forensics based on convolutional neural networks, IEEE Sig. Pro. Lett., 22 (2015), 1849–1853. |
| [16] | Y. Qian, J. Dong, W. Wang, et al., Deep learning for steganalysis via convolutional neural networks, Pro. SPIE. ISOE, 94 (2015), 9–14. |
| [17] | B. Belhassen and M. C. Stamm, A deep learning approach to universal image manipulation detection using a new convolutional layer, IH&MMSec, (2016), 5–10. |
| [18] | R. Yuan and J. Ni, A deep learning approach to detection of splicing and copy-move forgeries in images, WIFS, (2016), 1–6. |
| [19] | P. Zhou, X. Han, V. I. Morariu and L. S. Davis, Learning rich features for image manipulation detection,IEEE Conf. Comput. Vis. Pattern Recognit., Salt Lake, (2018), 1053–1061. |
| [20] | K. He, G. Gkioxari, P. Dollár, et al., Mask R-CNN. ICCV, 99 (2017), 1–11. |
| [21] | R. S. Zimmermann and J. N. J. a. p. a. Siems, Faster Training of Mask R-CNN by Focusing on Instance Boundaries, arXiv: 1809.07069. |
| [22] | C. Lopez-Molina, H. Bustince, J. Fernández, et al., A t-norm based approach to edge detection, IWCANN, Springer, Berlin, Heidelberg, (2009), 302–309. |
| [23] | T.Y. Lin, M. Maire, S. Belongie, et al., Microsoft coco: Common objects in context, ECCV, Springer, Cham, (2014), 740–755. |
| [24] | B. Wen, Y. Zhu, R. Subramanian, et al., Coverage novel database for copy-move forgery detection. ICIP, (2016), 161–165. |
| [25] | Y. F. Hsu and S. F. Chang, Detecting image splicing using geometry invariants and camera characteristics consistency, ICME, (2006), 549–552. |
| [26] | L. Jonathan, E. Shelhamer and T. Darrell, Fully Convolutional Networks for Semantic Segmentation, CVPR, 39 (2015), 3431–3440. |
| [27] | T.Y. Lin, P. Dollar, R. Girshick, et al., Feature pyramid networks for object detection, CVPR, (2017), 2117–2125. |
| [28] | N. Krawetz and H. F. J. H. F. S. Solutions, A Picture's Worth, Hacker Fact. Solut., 6 (2007), 1–31. |
| [29] | B. Mahdian and S. Saic, Using noise inconsistencies for blind image forensics, Image Vis. Comput., 7 (2009), 1497–1503. |
| [30] | P. Ferrara, T. Bianchi, A. De Rosa, et al., Image forgery localization via fine-grained analysis of CFA artifacts, IEEE Trans. Inf. Foren. Secur., 7(2012), 1566–1577. |
| [31] | J. H. Bappy, A. K. Roy-Chowdhury, J. Bunk, et al., Exploiting spatial structure for localizing manipulated image regions, ICCV, (2017), 4970–4979. |
| [32] | R. Salloum, Y. Ren and C.-C. J. Kuo, Image splicing localization using a multi-task fully convolutional network (MFCN), J. Vis. Com. Image Repre., 51 (2018), 201–209. |
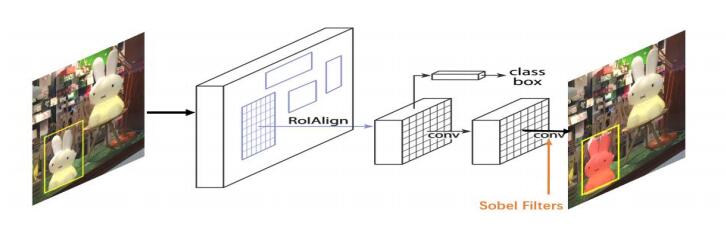
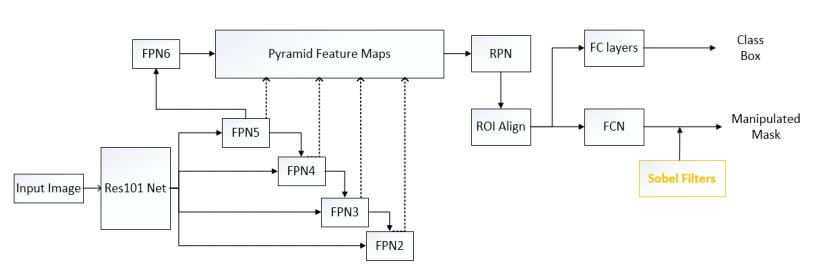
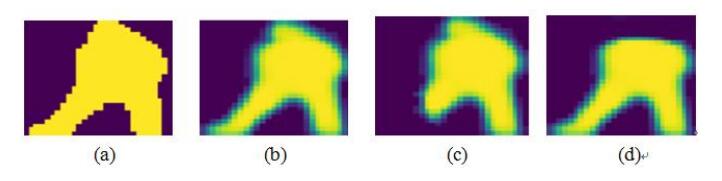
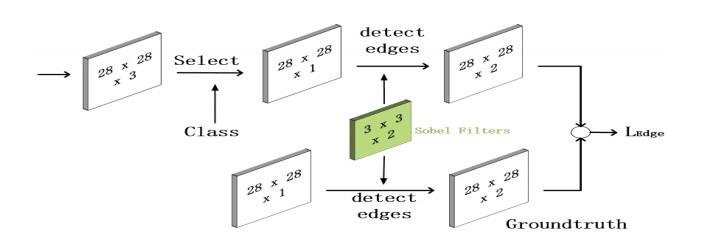


Figures(6) / Tables(5)

Xinyi Wang, He Wang, Shaozhang Niu, Jiwei Zhang. Detection and localization of image forgeries using improved mask regional convolutional neural network[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4581-4593. doi: 10.3934/mbe.2019229







 DownLoad:
DownLoad: