Citation: Xin-You Meng, Yu-Qian Wu. Bifurcation analysis in a singular Beddington-DeAngelis predator-prey model with two delays and nonlinear predator harvesting[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2668-2696. doi: 10.3934/mbe.2019133

| [1] | M. Kot, Elements of Mathematical Biology, Cambridge University Press, Cambridge, 2001. |
| [2] | S. Levin, T. Hallam and J. Cross, Applied Mathematical Ecology, Springer, New York, 1990. |
| [3] | R. May, Stability and Complexity in Model Ecosystems, Princeton University Press, Princeton, 1993. |
| [4] | C. Ji, D. Jing and N. Shi, A note on a predator-prey model with modified Leslie-Gower and Holling-type II schemes with stochastic perturbatio, J. Math. Anal. Appl., 377 (2011), 435–440. |
| [5] | T. Kar and H. Matsuda, Global dynamics and controllability of a harvested prey-predator system with Holling type III functional response, Nonlinear Anal. Hybrid Syst., 1 (2007), 59–67. |
| [6] | X. Meng, J. Wang, H. Huo, Dynamical behaviour of a nutrient-plankton model with Holling type IV, delay, and harvesting, Discrete Dyn. Nat. Soc., 2018 (2018), Article ID 9232590. |
| [7] | X. Meng, H. Huo, H. Xiang, et al., Stability in a predator-prey model with Crowley-Martin function and stage structure for prey, Appl. Math. Comput., 232 (2014), 810–819. |
| [8] | W. Yang, Diffusion has no influence on the global asymptotical stability of the Lotka-Volterra pre-predator model incorporating a constant number of prey refuges, Appl. Math. Comput., 223 (2013), 278–280. |
| [9] | Y. Zhu and K.Wang, Existence and global attractivity of positive periodic solutions for a predatorprey model with modified Leslie-Gower Holling-type II schemes, J. Math. Anal. Appl., 384 (2011), 400–408. |
| [10] | J. Liu, Dynamical analysis of a delayed predator-prey system with modified Leslie-Gower and Beddington-DeAngelis functional response, Adv. Difference Equ., 2014 (2014), 314–343. |
| [11] | X. Liu and Y. Wei, Dynamics of a stochastic cooperative predator-prey system with Beddington- DeAngelis functional response, Adv. Difference Equ., 2016 (2016), 21–39. |
| [12] | C. Li, X. Guo and D. He, An impulsive diffusion predator-prey system in three-species with Beddington-DeAngelis response, J. Appl. Math. Comput., 43 (2013), 235–248. |
| [13] | T. Ivanov and N. Dimitrova, A predator-prey model with generic birth and death rates for the predator and Beddington-DeAngelis functional response, Math. Comput. Simulat., 133 (2017), 111–123. |
| [14] | Q. Meng and L. Yang, Steady state in a cross-diffusion predator-prey model with the Beddington- DeAngelis functional response, Nonlinear Anal.: Real World Appl., 45 (2019), 401–413. |
| [15] | W. Liu, C. Fu and B. Chen, Hopf bifurcation and center stability for a predator-prey biological economic model with prey harvesting, Commun. Nonlinear Sci. Numer. Simul., 17 (2012), 3989– 3998. |
| [16] | F. Conforto, L. Desvillettes and C. Soresina, About reaction-diffusion systems involving the Holling-type II and the Beddington-DeAngelis functional responses for predator-prey models, Nonlinear Differ. Equ. Appl., 25 (2018), 24. |
| [17] | X. Sun, R. Yuan and L. Wang, Bifurcations in a diffusive predator-prey model with Beddington- DeAngelis functional response and nonselective harvesting, J. Nonlinear Sci., 29 (2019), 287–318. |
| [18] | J. Beddington, Mutual interference between parasites or predators and its effect on searching efficiency, J. Anim. Ecol., 44 (1975), 331–340. |
| [19] | D. DeAngilis, R. Goldstein and R. Neill, A model for tropic interaction, Ecology, 56 (1975), 881–892. |
| [20] | H. Freedman, Deterministic Mathematical Models in Population Ecology, Marcel Dekker, New York, 1980. |
| [21] | G. Seifert, Asymptotical behavior in a three-component food chain model, Nonlinear Anal. Theory Methods Appl., 32 (1998), 749–753. |
| [22] | C. Liu, Q. Zhang, X. Zhang, et al., Dynamical behavior in a stage-structured differential-algebraic prey-predator model with discrete time delay and harvesting, J. Comput. Appl. Math., 231 (2009), 612–625. |
| [23] | X. Meng, H. Huo, X. Zhang, et al., Stability and hopf bifurcation in a three-species system with feedback delays, Nonlinear Dyn., 64 (2011), 349–364. |
| [24] | X. Meng and Y. Wu, Bifurcation and control in a singular phytoplankton-zooplankton-fish model with nonlinear fish harvesting and taxation, Int. J. Bifurcat. Chaos, 28 (2018), 1850042(24 pages). |
| [25] | H. Xiang, Y. Wang and H. Huo, Analysis of the binge drinking models with demographics and nonlinear infectivity on networks, J. Appl. Anal. Comput., 8 (2018), 1535–1554. |
| [26] | K. Chakraborty, M. Chakraboty and T. Kar, Bifurcation and control of a bioeconomic model of a prey-predator system with a time delay, Nonlinear Anal. Hybrid Syst., 5 (2011), 613–625. |
| [27] | W. Liu, C. Fu and B. Chen, Hopf birfucation for a predator-prey biological economic system with Holling type II functional response, J. Franklin Inst., 348 (2011), 1114–1127. |
| [28] | G. Zhang, B. Chen, L. Zhu, et al., Hopf bifurcation for a differential-algebraic biological economic system with time delay, Appl. Math. Comput, 218 (2012), 7717–7726. |
| [29] | T. Faria, Stability and bifurcation for a delayed predator-prey model and the effect of diffusion, J. Math. Anal. Appl., 254 (2001), 433–463. |
| [30] | G. Zhang, Y. She and B. Chen, Hopf bifurcation of a predator prey system with predator harvesting and two delays, Nonlinear Dyn., 73 (2013), 2119–2131. |
| [31] | J. Zhang, Z. Jin, J. Yan, et al., Stability and Hopf bifurcation in a delayed competition system, Nonlinear Anal. Theory Methods Appl., 70 (2009), 658–670. |
| [32] | Z. Lajmiri, R. K. Ghaziani and I. Orak, Bifurcation and stability analysis of a ratio-dependent predator-prey model with predator harvesting rate, Chaos Soliton Fract., 106 (2018), 193–200. |
| [33] | M. Liu, X. He and J. Yu, Dynamics of a stochastic regime-switching predator-prey model with harvesting and distributed delays, Nonlinear Anal. Hybrid Syst., 28 (2018), 87–104. |
| [34] | T. Das, R. Mukerjee and K. Chaudhuri, Harvesting of a prey-predator fishery in the presence of toxicity, Appl. Math. Model., 33 (2009), 2282–2292. |
| [35] | R. Gupta and P. Chandra, Bifurcation analysis of modied Leslie-Gower predator-prey model with Michaelis-Menten type prey harvesting, J. Math. Anal. Appl., 398 (2013), 278–295. |
| [36] | P. Srinivasu, Bioeconomics of a renewable resource in presence of a predator, Nonlinear Anal. Real World Appl., 2 (2001), 497–506. |
| [37] | G. Lan, Y. Fu, C. Wei, et al., Dynamical analysis of a ratio-dependent predator-prey model with Holling III type functional response and nonlinear harvesting in a random environment, Adv. Differ. Equ., 2018 (2018), 198. |
| [38] | R. Gupta and P. Chandra, Dynamical complexity of a prey-predator model with nonlinear predator harvesting, Discrete Contin. Dyn. Syst. Ser. B, 20 (2015), 423–443. |
| [39] | J. Liu and L. Zhang, Bifurcation analysis in a prey-predator model with nonlinear predator harvesting, J. Franklin Inst., 353 (2016), 4701–4714. |
| [40] | K. Chakraborty, S. Jana and T. Kar, Global dynamics and bifurcation in a stage structured preypredator fishery model with harvesting, Appl. Math. Comput., 218 (2012), 9271–9290. |
| [41] | H. Gordon, The economic theory of a common property resource: the fishery, Bull. Math. Biol., 62 (1954), 124–142. |
| [42] | C. Liu, Q. Zhang and X. Duan, Dynamical behavior in a harvested differential-algebraic preypredator model with discrete time delay and stage structure, J. Franklin Inst., 346 (2009), 1038– 1059. |
| [43] | X. Zhang and Q. Zhang, Bifurcation analysis and control of a class of hybrid biological economic models, Nonlinear Anal. Hybrid Syst., 3 (2009), 578–587. |
| [44] | C. Liu, N. Lu, Q. Zhang, et al., Modelling and analysis in a prey-predator system with commercial harvesting and double time delays, J. Appl. Math. Comput., 281 (2016), 77–101. |
| [45] | M. Li, B. Chen and H. Ye, A bioeconomic differential algebraic predator-prey model with nonlinear prey harvesting, Appl. Math. Model., 42 (2017), 17–28. |
| [46] | P. Leslie and J. Gower, The properties of a stochastic model for the predator prey type of interaction between two species, Biometrika, 47 (1960), 219–234. |
| [47] | R. Cantrell and C. Cosner, On the dynamics of predator-prey models with the Beddington- DeAngelis functional response, J. Math. Anal. Appl., 257 (2001), 206–222. |
| [48] | L. Dai, Singular Control System, Springer, New York, 1989. |
| [49] | V. Venkatasubramanian, H. Schattler and J. Zaborszky, Local bifurcations and feasibility regions in differential-algebraic systems, IEEE Trans. Automat. Control, 40 (1995), 1992–2013. |
| [50] | Q. Zhang, C. Liu and X. Zhang, A singular bioeconomic model with diffusion and time delay, J. Syst. Sci. Complex., 24 (2011), 277–190. |
| [51] | J. Hale, Theory of Functional Differential Equations, Springer, New York, 1997. |
| [52] | B. Hassard, N. Kazarinoff and Y. Wan, Theory and Applications of Hopf Bifurcation, Cambridge University Press, Cambridge, 1981. |
| [53] | J. Guckenheimer and P. Holmes, Nonlinear Oscillations, Dynamical Systems, and Bifurcations of Vector Fields, Springer, New York, 1983. |
| [54] | Y. Kuang, Delay Differential Equations with Applications in Population Dynamics, Academic Press, New York, 1993. |
| [55] | H. Freedman and V. S. H. Rao, The trade-off between mutual interference and time lags in predator-prey systems, Bull. Math. Biol., 45 (1983), 991–1004. |
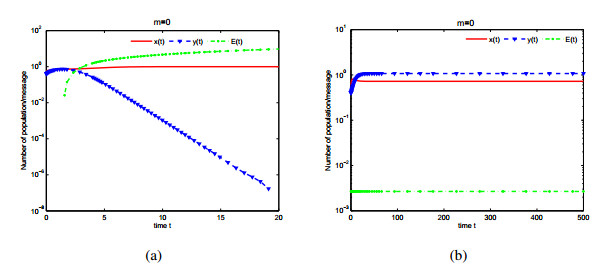
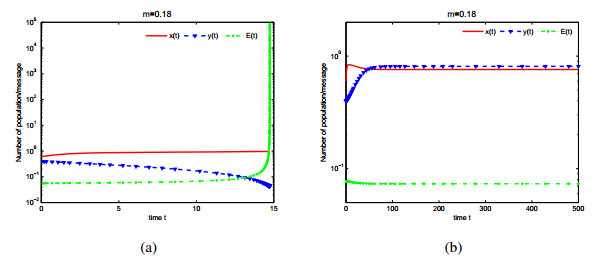
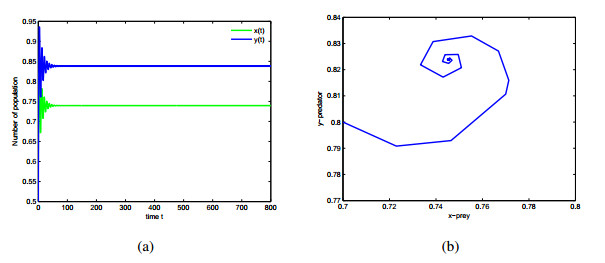
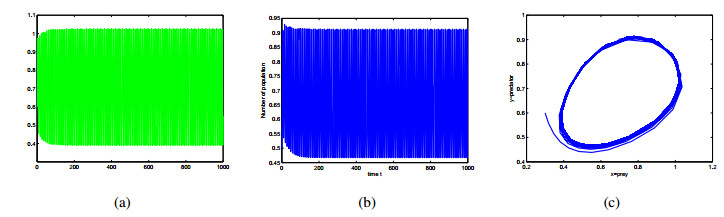
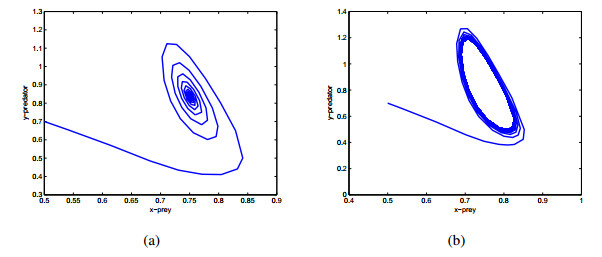
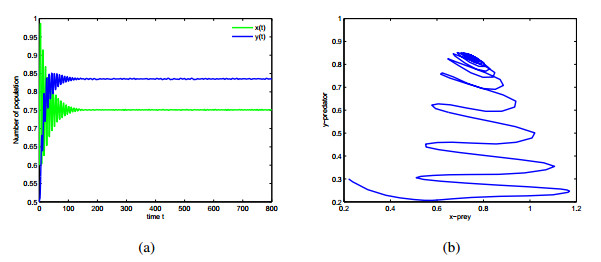
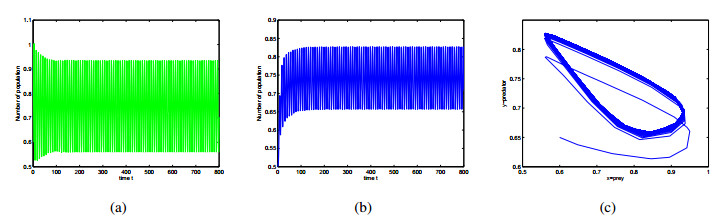
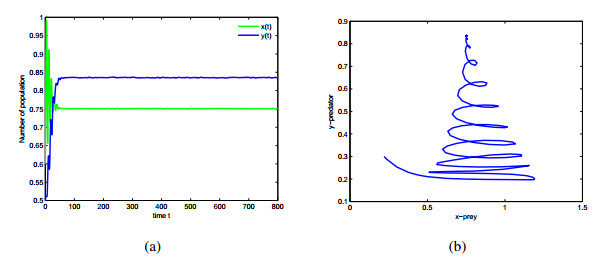
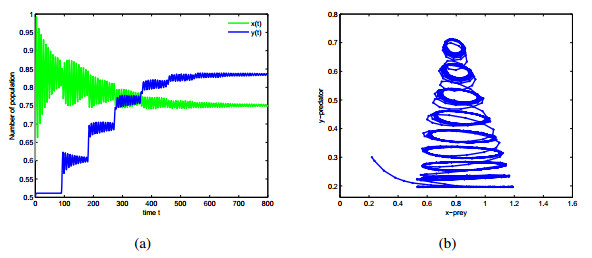
Figures(9)

Xin-You Meng, Yu-Qian Wu. Bifurcation analysis in a singular Beddington-DeAngelis predator-prey model with two delays and nonlinear predator harvesting[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2668-2696. doi: 10.3934/mbe.2019133







 DownLoad:
DownLoad: