Citation: Shuang-Hong Ma, Hai-Feng Huo. Global dynamics for a multi-group alcoholism model with public health education and alcoholism age[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1683-1708. doi: 10.3934/mbe.2019080

| [1] | J. Rehm, C. Mathers and S. Popova, et al., Global burden of disease and injury and economic cost attributable to alcohol use and alcohol-use disorders, Lancet, 373 (2009), 2223–2233. |
| [2] | J. Rehm, The risk associated with alcohol use and alcoholism, Alcohol Res. Health, 34 (2011), 135–143. |
| [3] | C. Lin, C. Lin and L. Mo, et al., Heavy alcohol consumption increases the incidence of hepatocellular carcinoma in hepatitis B virus-related cirrhosis, J. Hepatol, 58 (2013), 730–735. |
| [4] | WHO, Global Status Report on Alcohol and Health 2014, Geneva, Switzerland, 2014. |
| [5] | J. S. Gill, Reported levels of alcohol consumption and binge drinking within the UK undergraduate student population over the last 25 years, Alcohol, 37 (2002), 109–120. |
| [6] | L. Smith and D. Foxcroft, Drinking in the UK: An exploration of trends, 2009. |
| [7] | O. Sharomi and A. B. Gumel, Curtailing smoking dynamics: a mathematical modeling approach, Appl. Math. Comput., 195 (2008), 475–499. |
| [8] | H. F. Huo and X. M. Zhang, Complex dynamics in an alcoholism model with the impact of Twitter, Math. Biosci., 281 (2016), 24–35. |
| [9] | E. White and C. Comiskey, Heroin epidemics, treatment and ODE modelling, Math. Biosci., 208 (2007), 312–324. |
| [10] | G. Mulone and B. Straughan, A note on heroin epidemics, Math. Biosci., 218 (2009), 138-141. |
| [11] | B. Benedict, Modelling alcoholism as a contagious disease: how "infected" drinking buddies spread problem drinking, SIAM News, 40 2007. |
| [12] | J. L. Manthey, A. Aidoob and K. Y. Ward, Campus drinking: |
| [13] | an epidemiological model, J. Biol. Dyn., 2 (2008), 346–356. |
| [14] | 13 A. Mubayi, P. Greenwood and C. Castillo-Chavez, et al., The impact of relative residence times on the distribution of heavy drinkers in highly distinct environments, Socio. Econ. Plan Sci., 44 (2010), 45–56. |
| [15] | 14 R. Bani, R. Hameed and S. Szymanowski, et al., Influence of environmental factors on college alcohol drinking patterns, Math. Biosci. Eng., 10 (2013), 1281–1300. |
| [16] | 15 B. Buonomo and D. Lacitignola, Modeling peer influence effects on the spread of high-risk alcohol consumption behavior, Ricerche mat., 63 (2014), 101–117. |
| [17] | 16 A. Mubayi and P. Greenwood, Contextual interventions for controlling alcohol drinking, Math. Popul. Stud., 20 (2013), 27–53. |
| [18] | 17 G. Mulone and B. Straughan, Modeling binge drinking, Int. J. Biomath., 5 (2012), 57-70. |
| [19] | 18 H. F. Huo and N. N. Song, Golobal stability for a binge drinking model with two stages, Discrete Dyn. Nat. Soc., 2012, Article ID 829386, (2012), 15 pages. |
| [20] | 19 C. P. Bhunu and S. Mushayabasa, A theoretical analysis of smoking and alcoholism, J. Math. Model. Algor., 11 (2), 387–408. |
| [21] | 20 C. E. Walters, B. Straughan and R. Kendal, Modeling alcohol problems: total recovery, Ricerche Mat., 62 (2013), 33-53. |
| [22] | 21 H. Xiang, C. C. Zhu and H. F. Huo, Modelling the effect of immigration on drinking behaviour, J. Biol. Dyn., 11 (2017), 275–298. |
| [23] | 22 X. Y. Wang, H. F. Huo and Q. K. Kong, et al., Optimal control strategies in an alcoholism model, Abstr. Appl. Anal., 3 (2014), 1-18. |
| [24] | 23 C. C. Zhu and J. Zhu, Stability of a reaction-diffusion alcohol model with the impact of tax policy, Comput. Math. Appl., 74 (2017), 613–633. |
| [25] | 24 H. F. Huo, R. Chen and X. Y. Wang, Modelling and stability of HIV/AIDS epidemic model with treatment, Appl. Math. Model., 40 (2016), 6550–6559. |
| [26] | 25 H. F. Huo and M. X. Zou, Modelling effects of treatment at home on tuberculosis transmission dynamics, Appl. Math. Model., 40 (2016), 9474–9484. |
| [27] | 26 H. Xiang, Y. L. Tang and H. F. Huo, A viral model with intracellular delay and humoral immunity, Bull. Malays. Math. Sci. Soc., 40 (2017), 1011–1023. |
| [28] | 27 H. Xiang, Y. Y. Wang and H. F. Huo, Analysis of the binge drinking models with demographics and nonlinear infectivity on networks, J. Appl. Anal. Comput., 8 (2018), 1535–1554. |
| [29] | 28 Y. L. Cai, J. J. Jiao and Z. J. Gui, et al., Environmental variability in a stochastic epidemic model, Appl. Math. Comput., 3(2018), 210–226. |
| [30] | 29 Z. Du and Z. Feng, Existence and asymptotic behaviors of traveling waves of a modified vectordisease model, Commun. Pur. Appl. Anal., 17 (2018), 1899–1920. |
| [31] | 30 X. B. Zhang, Q. H. Shi and S. H. Ma, et al., Dynamic behavior of a stochastic SIQS epidemic model with levy jumps, Nonlinear Dynam., 93 (2018), 1481–1493. |
| [32] | 31 W. M. Wang, Y. L. Cai and Z. Q. Ding, et al., A stochastic differential equation SIS epidemic model incorporating Ornstein-Uhlenbeck process, Phys. A, 509 (2018), 921–936. |
| [33] | 32 H. F. Huo, F. F. Cui and H. Xiang, Dynamics of an SAITS alcoholism model on unweighted and weighted networks, Phys. A, 496 (2018), 249–262. |
| [34] | 33 X. Y. Meng and Y. Q. Wu, Bifurcation and control in a singular phytoplankton-zooplankton-fish model with nonlinear fish harvesting and taxation, Int. J. Bifurcat. Chaos, 28 (2018),1850042, 24 pages. |
| [35] | 34 A. K. Misra, A. Sharma and J. B. Shukla, Modeling and analysis of effects of awareness programs by media on spread of infectious diseases, Math. Comput. Model., 53 (2011), 1221–1228. |
| [36] | 35 H. F. Huo and Q. Wang, Modeling the influence of awareness programs by media on the drinking dynamics, Abstr. Appl. Anal., 2014 (2014), 1–8. |
| [37] | 36 H. Xiang, N. N. Song and H. F. Huo, Modelling effectss of public health educational campaigns on drinking dynamics, J. Biol. Dyn., 10 (2016), 164–178. |
| [38] | 37 S. H. Ma, H. F. Huo and X. Y. Meng, Modelling alcoholism as a contagious disease: A mathematical model with awareness programs and time delay, Discrete Dyn. Nat. Soc., 2015 (2015), 1–13. |
| [39] | 38 H. Guo, M. Y. Li and Z. Shuai, Global stability of the endemic equilibrium of multigroup SIR epidemic models, Can. Appl. Math. Q., 14 (2006), 259–284. |
| [40] | 39 H. Guo, M. Y. Li and Z. Shuai, A graph-theoretic approach to the method of global Lyapunov functions, Proc. Amer. Math. Soc., 136 (2008), 2793–2802. |
| [41] | 40 M. Y. Li and Z. Shuai, Global-stability problem for coupled systems of differential equations on networks, J. Diff. Equs., 248 (2010), 1–20. |
| [42] | 41 A. Berman and R. J. Plemmons, Nonnegative Matrices in the Mathematical Sciences, Academic Press, NewYork, 1979. |
| [43] | 42 Y. Muroya, Y. Enatsu and T. Kuniya, Global stability for a multi-group SIRS epidemic model with varying population sizes, Nonlinear Anal. RWA, 14 (2013), 1693–1704. |
| [44] | 43 R. Sun, Global stability of the endemic equilibrium of multigroup SIR models with nonlinear incidence, Comput. Math. Appl., 60 (2010), 2286–2291. |
| [45] | 44 M. Y. Li, Z. Shuai and C. Wang, Global stability of multi-group epidemic models with distributed delays, J. Math. Anal. Appl., 361 (2010), 38–47. |
| [46] | 45 Y. Muroya and T. Kuniya, Further stability analysis for a multi-group SIRS epidemic model with varying total population size, Appl. Math. Lett., 38 (2014), 73–78. |
| [47] | 46 J. Wang and H. Shu, Global dynamics of a multi-group epidemic model with latency, relapse and nonlinear incidence rate, Math. Biosci. Eng., 13 (2016), 209–225. |
| [48] | 47 F. Hoppensteadt, An age-dependent epidemic model, J. Franklin Inst., 297 (1974), 325–338. |
| [49] | 48 X. Z. Li, J. Wang and M. Ghosh, Stability and bifurcation of an SIVS epidemic model with treatment and age of vaccination, Appl. Math. Model., 34 (2010), 437–450. |
| [50] | 49 X. C. Duan, S. L. Yuan and X. Z. Li, Global stability of an SVIR model with age of vaccination, Appl. Math. Comput., 226 (2014), 528–540. |
| [51] | 50 M. W. Shen and Y. N. Xiao, Global stability of a multi-group SVEIR epidemiological model with the vaccination age and infection age, Acta Appl. Math., 144 (2016), 137–157. |
| [52] | 51 J. H. Xu and Y. C. Zhou, Global stability of a multi-group model with generalized nonlinear incidence and vaccination age, Discrete Contin. Dyn. Syst. Ser. B, 21 (2016), 977–996. |
| [53] | 52 F. V. Atkinson and J. R. Haddock, On determining phase spaces for functional differential equations, Funkc. Ekvacioj., 31 (1988), 331–347. |
| [54] | 53 G. F.Webb, Theory of Nonlinear Age-dependent Population Dynamics, Marcel Dekker, New York, 1985. |
| [55] | 54 G. Gripenberg, S. O. Londen and O. Staffans, Volterra Integral and Functional Equations, Cambridge University Press, New York, 1990. |
| [56] | 55 O. Diekmann, J. Heesterbeek and J. A. Metz, On the defnition and the computation of the basic reproduction ratio R0 in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28 (1990), 365–382. |
| [57] | 56 P. van den Driessche and J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
| [58] | 57 J. K. Hale and P. Waltman, Presistence in infinite-dimensional systems, SIAM J. Appl. Math., 20 (1989), 388–395. |
| [59] | 58 A. Berman and R. J. Plemmons, Nonnegative Matrices in the Mathematical Sciences, Academic Press, NewYork, 1979. |
| [60] | 59 J. P. Lasalle, The Stability of Dynamical Systems, in: Regional Conference Series in Applied Mathematics, SIAM, Philadelphia, 1976. |
| [61] | 60 J. K. Hale and S .M. V. Lunel, Introduction to Functional Differential Equations, Appl. Math. Sci., 99, Springer, New York, 1993. |
| [62] | 61 H. I. Freedman, S. Ruan and M. Tang, Uniform persistence and flows near a closed positively invariant set, J. Dyn. Diff. Equat., 6 (1994), 583–600. |
| [63] | 62 M. Y. Li, J. R. Graef and L. Wang, et al., Global dynamics of a SEIR model with varying total population size, Math. Biosci., 160 (1999), 191–213. |
| [64] | 63 N. P. Bhatia and G. P. Szego, Dynamical Systems: Stability Theory and Applications, Lecture Notes in Mathematics, 35, Springer, Berlin, 1967. |
| [65] | 64 H. L. Smith and P. Waltman,The Theory of the Chemostat: Dynamics of Microbial Competition, Cambridge University Press, Cambridge, 1995. |
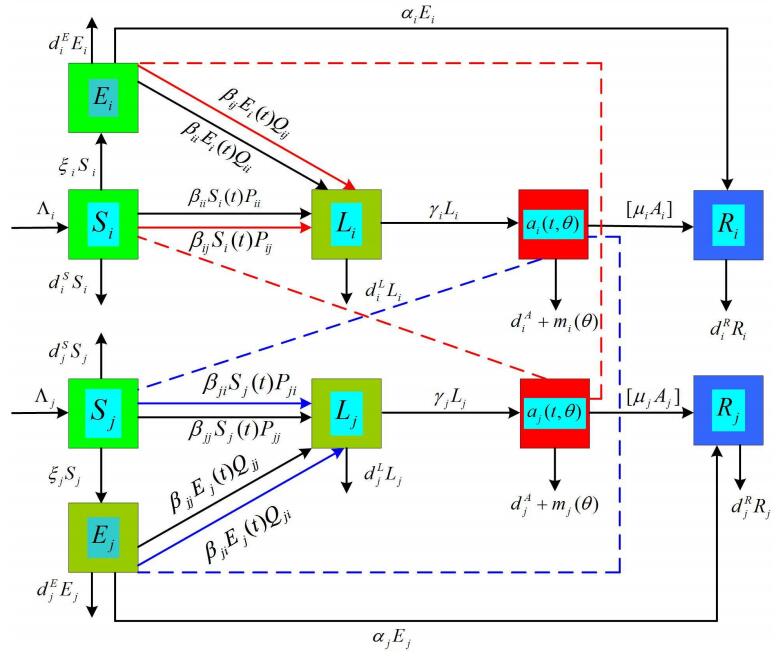
Figures(1) / Tables(1)

Shuang-Hong Ma, Hai-Feng Huo. Global dynamics for a multi-group alcoholism model with public health education and alcoholism age[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1683-1708. doi: 10.3934/mbe.2019080







 DownLoad:
DownLoad: