Citation: Conrad Ratchford, Jin Wang. Modeling cholera dynamics at multiple scales: environmental evolution, between-host transmission, and within-host interaction[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 782-812. doi: 10.3934/mbe.2019037

| [1] | J. R. Andrews and S. Basu, Transmission dynamics and control of cholera in Haiti: an epidemic model, Lancet, 377 (2011), 1248–1255. |
| [2] | A. Bechette, T. Stojsavljevic, M. Tessmer, J. A. Berges, G. A. Pinter and E. B. Young, Mathematical modeling of bacteria-virus interactions in Lake Michigan incorporating phosphorus content,J. Great Lakes Res., 39 (2013), 646–654. |
| [3] | B. Boldin and O. Diekmann, Superinfections can induce evolutionarily stable coexistence of pathogens,J. Math. Biol., 56 (2008), 635–672. |
| [4] | V. Capasso and S. L. Paveri-Fontana, A mathematical model for the 1973 cholera epidemic in the European Mediterranean region,Rev. Epidemiol. Sante, 27 (1979), 121–132. |
| [5] | C. Castillo-Chavez, Z. Feng and W. Huang, On the computation of R0 and its role in global stability, inMathematical Approaches for Emerging and Reemerging Infectious Diseases: an Introduction, (2002), 229–250. |
| [6] | C. Castillo-Chavez and B. Song, Dynamical models of tuberculosis and their applications,Math. Biosci. Eng., 1 (2004), 361–404. |
| [7] | R. R. Colwell, A global and historical perspective of the genus Vibrio, inThe Biology of Vibrios, (eds. F.L. Thompson, B. Austin and J. Swings), ASM Press: Washington DC, 2006. |
| [8] | F. R. Gantmacher and J. L. Brenner,Applications of the Theory of Matrices, Dover: Mineola, 2005. |
| [9] | D. M. Hartley, J. G. Morris and D. L. Smith, Hyperinfectivity: a critical element in the ability of V. cholerae to cause epidemics? PLoS Med., 3 (2006), 0063–0069. |
| [10] | H. W. Hethcote, The mathematics of infectious diseases,SIAM Rev., 42 (200) 599–653. |
| [11] | M. Y. Li and J. S. Muldowney, A geometric approach to global-stability problems,SIAM J. Math. Anal., 27 (1996), 1070–1083. |
| [12] | N. Mideo, S. Alizon and T. Day, Linking within- and between-host disease dynamics, Trends Ecol. Evol., 23 (2008), 511–517. |
| [13] | Z. Mukandavire, S. Liao, J. Wang, H. Gaff, D. L. Smith and J. G. Morris, Estimating the reproductive numbers for the 2008-2009 cholera outbreaks in Zimbabwe,Proc. Natl. Acad. Sci. USA, 108 (2011), 8767–8772. |
| [14] | J. D. Murray,Mathematical Biology, Springer, 2002. |
| [15] | E. J. Nelson, J. B. Harris, J. G. Morris, S. B. Calderwood and A. Camilli, Cholera transmission: The host, pathogen and bacteriophage dynamics,Nat. Rev. Microbiol., 7 (2009), 693–702. |
| [16] | L. Perko,Differential Equations and Dynamical Systems, Springer Science & Business Media: New York, 2013. |
| [17] | D. Posny and J. Wang, Modelling cholera in periodic environments,J. Biol. Dyn., 8 (2014) 1–19. |
| [18] | Z. Shuai and P. van den Driessche, Global stability of infectious disease models using Lyapunov functions,SIAM J. Appl. Math., 73 (2013), 1513–1532. |
| [19] | H. L. Smith and R. T. Trevino, Bacteriophage infection dynamics: multiple host binding sites, Math. Model. Nat. Pheno., 4 (2009), 111–136. |
| [20] | S.H. Strogatz,Nonlinear Dynamics and Chaos: with Applications to Physics, Biology, Chemistry, and Engineering, Westview Press: Boulder, 2015. |
| [21] | H. R. Thieme,Mathematics in Population Biology, Princeton University Press: Princeton, 2003. |
| [22] | J. P. Tian and J. Wang, Global stability for cholera epidemic models,Math. Biosci., 232 (2011), 31–41. |
| [23] | J. H. Tien and D. J. D. Earn, Multiple transmission pathways and disease dynamics in a waterborne pathogen model,Bull. Math. Biol., 72 (2010), 1506–1533. |
| [24] | P. van den Driessche and J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission,Math. Biosci., 180 (2002), 29–48. |
| [25] | M. K.Waldor and J. J. Mekalanos, Lysogenic conversion by a filamentous phage encoding cholera toxin,Science, 272 (1996), 1910–1914. |
| [26] | J. Wang and S. Liao, A generalized cholera model and epidemic-endemic analysis,J. Biol. Dyn., 6 (2012), 568–589. |
| [27] | X.Wang and J.Wang, Analysis of cholera epidemics with bacterial growth and spatial movement, J. Biol. Dyn., 9 (1) (2015), 233–261. |
| [28] | X. Wang, D. Gao and J. Wang, Influence of human behavior on cholera dynamics,Math. Biosci., 267 (2015), 41–52. |
| [29] | X. Wang and J. Wang, Disease dynamics in a coupled cholera model linking within-host and between-host interactions,J. Biol. Dyn., 11(S1) (2017), 238–262. |
| [30] | X. Wang and J. Wang, Modeling the within-host dynamics of cholera: bacterial-viral interaction, J. Biol. Dyn., 11(S2) (2017), 484–501. |
| [31] | WHO Weekly Epidemiology Bulletin, 2-8 April 2018. |


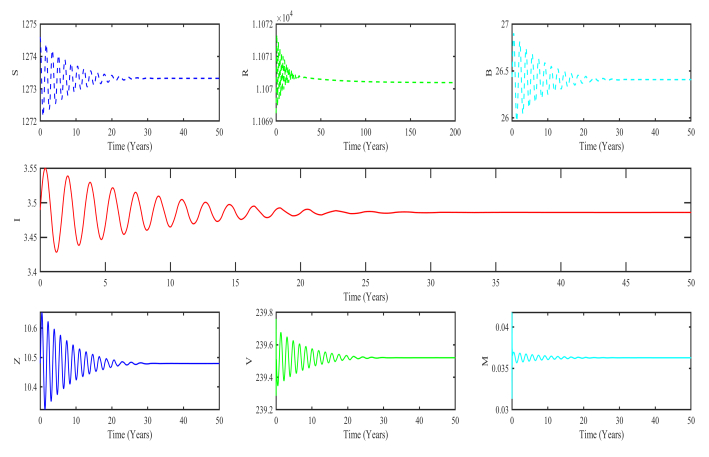
Figures(3)

Conrad Ratchford, Jin Wang. Modeling cholera dynamics at multiple scales: environmental evolution, between-host transmission, and within-host interaction[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 782-812. doi: 10.3934/mbe.2019037







 DownLoad:
DownLoad: