Citation: Raimund BÜrger, Gerardo Chowell, Elvis GavilÁn, Pep Mulet, Luis M. Villada. Numerical solution of a spatio-temporal gender-structured model for hantavirus infection in rodents[J]. Mathematical Biosciences and Engineering, 2018, 15(1): 95-123. doi: 10.3934/mbe.2018004

| [1] | [ G. Abramson and V. M. Kenkre, Spatiotemporal patterns in the Hantavirus infection Phys. Rev. E 66 (2002), 011912 (5pp). |
| [2] | [ M. A. Aguirre, G. Abramson, A. R. Bishop and V. M. Kenkre, Simulations in the mathematical modeling of the spread of the Hantavirus Phys. Rev. E 66 (2002), 041908 (5pp). |
| [3] | [ L. J. S. Allen,B. M. Bolker,Y. Lou,A. L. Nevai, Asymptotic of the steady states for an SIS epidemic patch model, SIAM J. Appl. Math., 67 (2007): 1283-1309. |
| [4] | [ L. J. S. Allen,R. K. McCormack,C. B. Jonsson, Mathematical models for hantavirus infection in rodents, Bull. Math. Biol., 68 (2006): 511-524. |
| [5] | [ R. M. Anderson,R. M. May, null, Infectious Diseases of Humans: Dynamics and Control, , Oxford Science Publications, 1991. |
| [6] | [ J. Arino, Diseases in metapopulations. In Z. Ma, Y. Zhou and J. Wu (Eds. ), Modeling and Dynamics of Infectious Diseases, Higher Education Press, Beijing, 11 (2009), 64-122. |
| [7] | [ J. Arino,J. R. Davis,D. Hartley,R. Jordan,J. M. Miller,P. van den Driessche, A multi-species epidemic model with spatial dynamics, Mathematical Medicine and Biology, 22 (2005): 129-142. |
| [8] | [ U. Ascher,S. Ruuth,J. Spiteri, Implicit-explicit Runge-Kutta methods for time dependent partial differential equations, Appl. Numer. Math., 25 (1997): 151-167. |
| [9] | [ P. Bi,X. Wu,F. Zhang,K. A. Parton,S. Tong, Seasonal rainfall variability, the incidence of hemorrhagic fever with renal syndrome, and prediction of the disease in low-lying areas of China, Amer. J. Epidemiol., 148 (1998): 276-281. |
| [10] | [ S. Boscarino,R. Bürger,P. Mulet,G. Russo,L. M. Villada, Linearly implicit IMEX Runge-Kutta methods for a class of degenerate convection-diffusion problems, SIAM J. Sci. Comput., 37 (2015): B305-B331. |
| [11] | [ S. Boscarino,F. Filbet,G. Russo, High order semi-implicit schemes for time dependent partial differential equations, J. Sci. Comput., 68 (2016): 975-1001. |
| [12] | [ S. Boscarino,P. G. LeFloch,G. Russo, High-order asymptotic-preserving methods for fully nonlinear relaxation problems, SIAM J. Sci. Comput., 36 (2014): A377-A395. |
| [13] | [ S. Boscarino,G. Russo, On a class of uniformly accurate IMEX Runge-Kutta schemes and applications to hyperbolic systems with relaxation, SIAM J. Sci. Comput., 31 (2009): 1926-1945. |
| [14] | [ S. Boscarino,G. Russo, Flux-explicit IMEX Runge-Kutta schemes for hyperbolic to parabolic relaxation problems, SIAM J. Numer. Anal., 51 (2013): 163-190. |
| [15] | [ F. Brauer,C. Castillo-Chavez, null, Mathematical Models in Population Biology and Epidemiology, Second Ed., Springer, New York, 2012. |
| [16] | [ M. Brummer-Korvenkontio,A. Vaheri,T. Hovi,C. H. von Bonsdorff,J. Vuorimies,T. Manni,K. Penttinen,N. Oker-Blom,J. Lähdevirta, Nephropathia epidemica: Detection of antigen in bank voles and serologic diagnosis of human infection, J. Infect. Dis., 141 (1980): 131-134. |
| [17] | [ J. Buceta, C. Escudero, F. J. de la Rubia and K. Lindenberg, Outbreaks of Hantavirus induced by seasonality Phys. Rev. E 69 (2004), 021908 (9pp). |
| [18] | [ R. Bürger,G. Chowell,P. Mulet,L. M. Villada, Modelling the spatial-temporal progression of the 2009 A/H1N1 influenza pandemic in Chile, Math. Biosci. Eng., 13 (2016): 43-65. |
| [19] | [ R. Bürger,R. Ruiz-Baier,C. Tian, Stability analysis and finite volume element discretization for delay-driven spatio-temporal patterns in a predator-prey model, Math. Comput. Simulation, 132 (2017): 28-52. |
| [20] | [ R. M. Colombo,E. Rossi, Hyperbolic predators versus parabolic preys, Commun. Math. Sci., 13 (2015): 369-400. |
| [21] | [ M. Crouzeix, Une méthode multipas implicite-explicite pour l'approximation des équations d'évolution paraboliques, Numer. Math., 35 (1980): 257-276. |
| [22] | [ O. Diekmann, H. Heesterbeek and T. Britton, Mathematical Tools for Understanding Infectious Disease Dynamics Princeton Series in Theoretical and Computational Biology, Princeton University Press, Princeton, NJ, 2013. |
| [23] | [ R. Donat,I. Higueras, On stability issues for IMEX schemes applied to 1D scalar hyperbolic equations with stiff reaction terms, Math. Comp., 80 (2011): 2097-2126. |
| [24] | [ C. Escudero, J. Buceta, F. J. de la Rubia and K. Lindenberg, Effects of internal fluctuations on the spreading of Hantavirus Phys. Rev. E 70 (2004), 061907 (7pp). |
| [25] | [ S. de Franciscis and A. d'Onofrio, Spatiotemporal bounded noises and transitions induced by them in solutions of the real Ginzburg-Landau model Phys. Rev. E 86 (2012), 021118 (9pp); Erratum, Phys. Rev. E 94 (2016), 0599005(E) (1p). |
| [26] | [ M. Garavello and B. Piccoli, Traffic Flow on Networks. Conservation Laws Models Amer. Inst. Math. Sci. , Springfield, MO, USA, 2006. |
| [27] | [ G. S. Jiang,C.-W. Shu, Efficient implementation of weighted ENO schemes, J. Comput. Phys., 126 (1996): 202-228. |
| [28] | [ P. Kachroo,S. J. Al-Nasur,S. A. Wadoo,A. Shende, null, Pedestrian Dynamics, , Springer-Verlag, Berlin, 2008. |
| [29] | [ A. Källén, Thresholds and travelling waves in an epidemic model for rabies, Nonlin. Anal. Theor. Meth. Appl., 8 (1984): 851-856. |
| [30] | [ A. Källén,P. Arcuri,J. D. Murray, A simple model for the spatial spread and control of rabies, J. Theor. Biol., 116 (1985): 377-393. |
| [31] | [ Y. Katznelson, null, An Introduction to Harmonic Analysis, Third Ed., Cambridge University Press, Cambridge, UK, 2004. |
| [32] | [ C. A. Kennedy,M. H. Carpenter, Additive Runge-Kutta schemes for convection-diffusion-reaction equations, Appl. Numer. Math., 44 (2003): 139-181. |
| [33] | [ W. O. Kermack,A. G. McKendrick, A contribution to the mathematical theory of epidemics, Proc. Roy. Soc. A, 115 (1927): 700-721. |
| [34] | [ N. Kumar, R. R. Parmenter and V. M. Kenkre, Extinction of refugia of hantavirus infection in a spatially heterogeneous environment Phys. Rev. E 82 (2010), 011920 (8pp). |
| [35] | [ T. Kuniya,Y. Muroya,Y. Enatsu, Threshold dynamics of an SIR epidemic model with hybrid and multigroup of patch structures, Math. Biosci. Eng., 11 (2014): 1375-1393. |
| [36] | [ H. N. Liu, L. D. Gao, G. Chowell, S. X. Hu, X. L. Lin, X. J. Li, G. H. Ma, R. Huang, H. S. Yang, H. Tian and H. Xiao, Time-specific ecologic niche models forecast the risk of hemorrhagic fever with renal syndrome in Dongting Lake district, China, 2005-2010, PLoS One, 9 (2014), e106839 (8pp). |
| [37] | [ X.-D. Liu,S. Osher,T. Chan, Weighted essentially non-oscillatory schemes, J. Comput. Phys., 115 (1994): 200-212. |
| [38] | [ H. Malchow,S. V. Petrovskii,E. Venturino, null, Spatial Patterns in Ecology and Epidemiology: Theory, Models, and Simulation, , Chapman & Hall/CRC, Boca Raton, FL, USA, 2008. |
| [39] | [ J. N. Mills,B. A. Ellis,K. T. McKee,J. I. Maiztegui,J. E. Childs, Habitat associations and relative densities of rodent populations in cultivated areas of central Argentina, J. Mammal., 72 (1991): 470-479. |
| [40] | [ P. A. P. Moran, Notes on continuous stochastic phenomena, Biometrika, 37 (1950): 17-23. |
| [41] | [ J. D. Murray, null, Mathematical Biology Ⅱ: Spatial Models and Biomedical Applications, Third Edition, Springer, New York, 2003. |
| [42] | [ J. D. Murray,E. A. Stanley,D. L. Brown, On the spatial spread of rabies among foxes, Proc. Roy. Soc. London B, 229 (1986): 111-150. |
| [43] | [ A. Okubo,S. A. Levin, null, Diffusion and Ecological Problems: Modern Perspectives, Second Edition, Springer-Verlag, New York, 2001. |
| [44] | [ O. Ovaskainen and E. E. Crone, Modeling animal movement with diffusion, in S. Cantrell, C. Cosner and S. Ruan (Eds. ), Spatial Ecology, Chapman & Hall/CRC, Boca Raton, FL, USA, 2009, 63-83. |
| [45] | [ L. Pareschi,G. Russo, Implicit-Explicit Runge-Kutta schemes and applications to hyperbolic systems with relaxation, J. Sci. Comput., 25 (2005): 129-155. |
| [46] | [ J. A. Reinoso and F. J. de la Rubia, Stage-dependent model for the Hantavirus infection: The effect of the initial infection-free period Phys. Rev. E 87 (2013), 042706 (6pp). |
| [47] | [ J. A. Reinoso and F. J. de la Rubia, Spatial spread of the Hantavirus infection Phys. Rev. E 91 (2015), 032703 (5pp). |
| [48] | [ R. Riquelme,M. L. Rioseco,L. Bastidas,D. Trincado,M. Riquelme,H. Loyola,F. Valdivieso, Hantavirus pulmonary syndrome, southern chile, 1995-2012, Emerg. Infect. Dis., 21 (2015): 562-568. |
| [49] | [ C. Robertson, C. Mazzetta and A. d'Onofrio, Regional variation and spatial correlation, Chapter 5 in P. Boyle and M. Smans (Eds. ), Atlas of Cancer Mortality in the European Union and the European Economic Area 1993-1997, IARC Scientific Publication, WHO Press, Geneva, Switzerland, 159 (2008), 91-113. |
| [50] | [ E. Rossi,V. Schleper, Convergence of a numerical scheme for a mixed hyperbolic-parabolic system in two space dimensions, ESAIM Math. Modelling Numer. Anal., 50 (2016): 475-497. |
| [51] | [ S. Ruan and J. Wu, Modeling spatial spread of communicable diseases involving animal hosts, in S. Cantrell, C. Cosner and S. Ruan (Eds. ), Spatial Ecology, Chapman & Hall/CRC, Boca Raton, FL, USA, 2010,293-316. |
| [52] | [ L. Sattenspiel, The Geographic Spread of Infectious Diseases: Models and Applications Princeton Series in Theoretical and Computational Biology, Princeton University Press, 2009. |
| [53] | [ C.-W. Shu,S. Osher, Efficient implementation of essentially non-oscillatory shock-capturing schemes, Ⅱ, J. Comput. Phys., 83 (1988): 32-78. |
| [54] | [ S. W. Smith, Digital Signal Processing: A Practical Guide for Engineers and Scientists. Demystifying technology series: by engineers, for engineers. Newnes, 2003. |
| [55] | [ H. Y. Tian, P. B. Yu, A. D. Luis, P. Bi, B. Cazelles, M. Laine, S. Q. Huang, C. F. Ma, S. Zhou, J. Wei, S. Li, X. L. Lu, J. H. Qu, J. H. Dong, S. L. Tong, J. J. Wang, B. Grenfell and B. Xu, Changes in rodent abundance and weather conditions potentially drive hemorrhagic fever with renal syndrome outbreaks in Xi'an, China, 2005-2012, PLoS Negl. Trop. Dis. , 9 (2015), paper e0003530 (13pp). |
| [56] | [ M. Treiber,A. Kesting, null, Traffic Flow Dynamics, , Springer-Verlag, Berlin, 2013. |
| [57] | [ P. van den Driessche, Deterministic compartmental models: Extensions of basic models, In F. Brauer, P. van den Driessche and J. Wu (Eds. ), Mathematical Epidemiology, SpringerVerlag, Berlin, 1945 (2008), 147-157. |
| [58] | [ P. van den Driessche, Spatial structure: Patch models, In F. Brauer, P. van den Driessche and J. Wu (Eds. ), Mathematical Epidemiology, Springer-Verlag, Berlin, 1945 (2008), 179-189. |
| [59] | [ P. van den Driessche,J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002): 29-48. |
| [60] | [ E. Vynnycky,R. E. White, null, An Introduction to Infectious Disease Modelling, , Oxford University Press, 2010. |
| [61] | [ J. Wu, Spatial structure: Partial differential equations models, In F. Brauer, P. van den Driessche and J. Wu (Eds. ), Mathematical Epidemiology, Springer-Verlag, Berlin, 2008,191-203. |
| [62] | [ H. Xiao,X. L. Lin,L. D. Gao,X. Y. Dai,X. G. He,B. Y. Chen, Environmental factors contributing to the spread of hemorrhagic fever with renal syndrome and potential risk areas prediction in midstream and downstream of the Xiangjiang River [in Chinese], Scientia Geographica Sinica, 33 (2013): 123-128. |
| [63] | [ C. J. Yahnke,P. L. Meserve,T. G. Ksiazek,J. N. Mills, Patterns of infection with Laguna Negra virus in wild populations of Calomys laucha in the central Paraguayan chaco, Am. J. Trop. Med. Hyg., 65 (2001): 768-776. |
| [64] | [ W. Y. Zhang,L. Q. Fang,J. F. Jiang,F. M. Hui,G. E. Glass,L. Yan,Y. F. Xu,W. J. Zhao,H. Yang,W. Liu, Predicting the risk of hantavirus infection in Beijing, People's Republic of China, Am. J. Trop. Med. Hyg., 80 (2010): 678-683. |
| [65] | [ W. Y. Zhang,W. D. Guo,L. Q. Fang,C. P. Li,P. Bi,G. E. Glass,J. F. Jiang,S. H. Sun,Q. Qian,W. Liu,L. Yan,H. Yang,S. L. Tong,W. C. Cao, Climate variability and hemorrhagic fever with renal syndrome transmission in Northeastern China, Environ. Health Perspect, 118 (2010): 915-920. |
| [66] | [ X. Zhong, Additive semi-implicit Runge-Kutta methods for computing high-speed nonequilibrium reactive flows, J. Comput. Phys., 128 (1996): 19-31. |
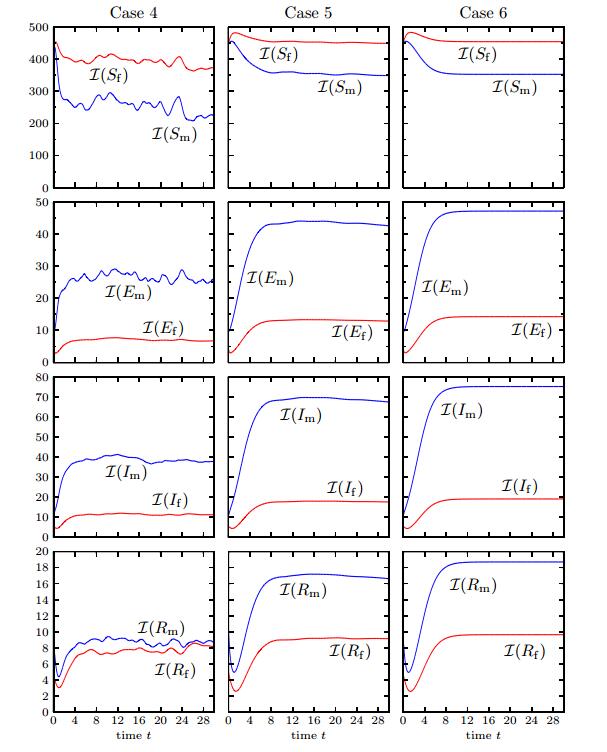
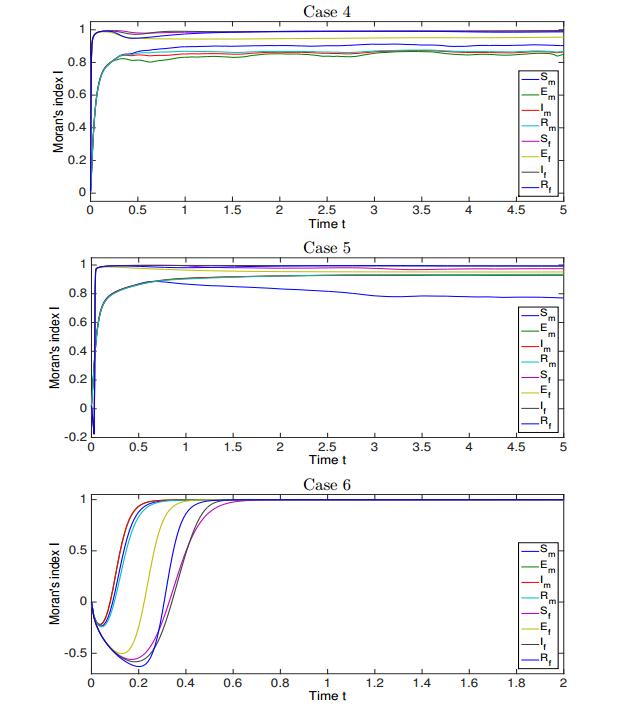
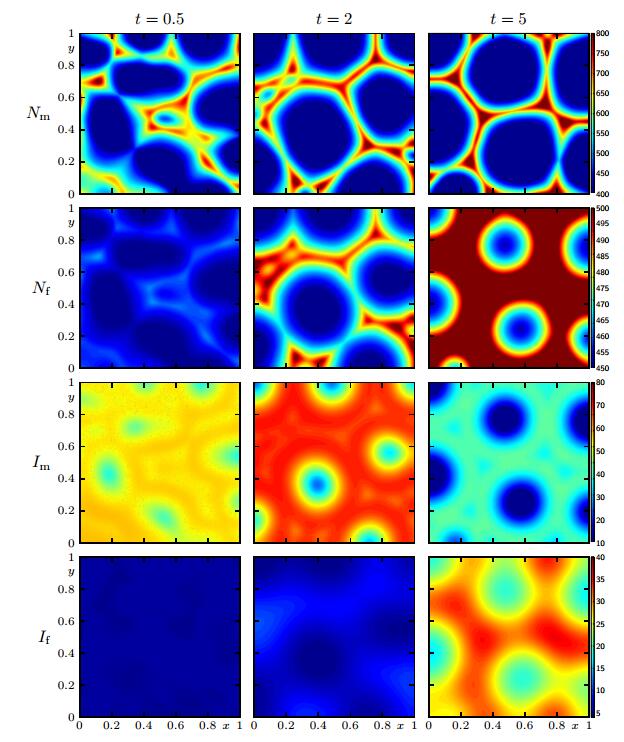
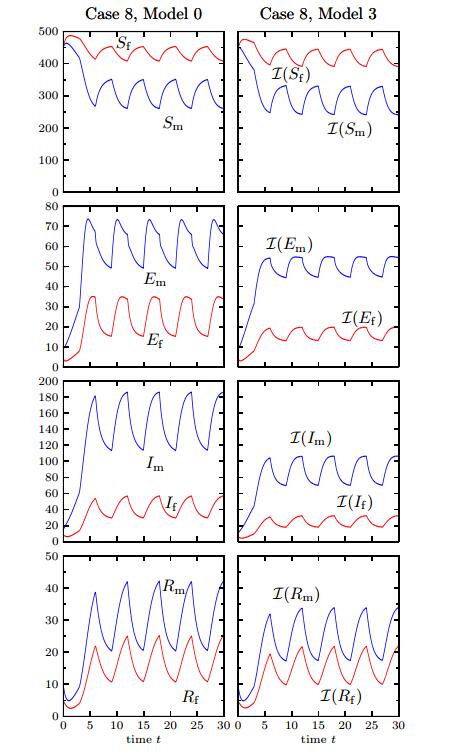
Figures(12) / Tables(1)

Raimund BÜrger, Gerardo Chowell, Elvis GavilÁn, Pep Mulet, Luis M. Villada. Numerical solution of a spatio-temporal gender-structured model for hantavirus infection in rodents[J]. Mathematical Biosciences and Engineering, 2018, 15(1): 95-123. doi: 10.3934/mbe.2018004







 DownLoad:
DownLoad: