Citation: Ming Chen, Meng Fan, Xing Yuan, Huaiping Zhu. Effect of seasonal changing temperature on the growth of phytoplankton[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1091-1117. doi: 10.3934/mbe.2017057

| [1] | [ G. Ahlgren, Temperature functions in biology and their application to algal growth constants, Oikos, 49 (1987): 177-190. |
| [2] | [ O. Bernard,B. Remond, Validation of a simple model accounting for light and temperature effect on microalgal growth, Bioresource Technology, 123 (2012): 520-527. |
| [3] | [ R. Bouterfas,M. Belkoura,A. Dauta, Light and temperature effects on the growth rate of three freshwater [2pt] algae isolated from a eutrophic lake, Hydrobiologia, 489 (2002): 207-217. |
| [4] | [ C. Butterwick,S.I. Heaney,J. F. Talling, Diversity in the influence of temperature on the growth rates of freshwater algae, and its ecological relevance, Freshwater Biol, 50 (2005): 291-300. |
| [5] | [ M. Chen,M. Fan,R. Liu,X. Yuan,H. P. Zhu, The dynamics of temperature and light on the growth of phytoplankton, J. Theor. Biol., 385 (2015): 8-19. |
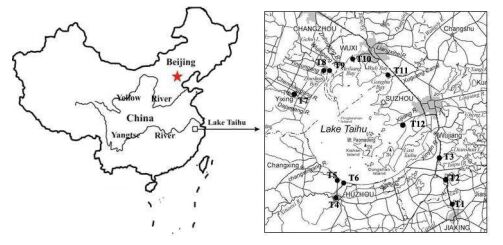
| [6] | [ W. Chen,A. Nauwerck, A note on composition and feeding of the crustacean zooplankton of Lake Taihu, Jiangsu Province, China, Limnologica, 26 (1996): 275-280. |
| [7] | [ Y. W. Chen,B. Q. Qin,K. Teubner,M. T. Dokulil, Long-term dynamics of phytoplankton assemblages: Microcystis-domination in Lake Taihu, a large shallow lake in China, J. Plankton Res., 25 (2003): 445-453. |
| [8] | [ L. Ding,Y. Pang,L. Li, Simulation study on algal dynamics under different hydrodynamic conditions, Acta Ecologica Sinica, 25 (2005): 1863-1868. |
| [9] | [ X. H. Dong,H. Bennion,R. Battarbee,X. D. Yang,E. F. Liu, Tracking eutrophication in Taihu Lake using the diatom record: potential and problems, J. Paleolimnol, 40 (2008): 413-429. |
| [10] | [ P. Driessche,W. James, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002): 29-48. |
| [11] | [ J. J. Elser,K. Acharya,M. Kyle,J. Cotner,W. Makino,T. Markow,T. Watts,S. Hobbie,W. Fagan,J. Schade,J. Hood,R. W. Sterner, Growth rate-stoichiometry couplings in diverse biota, Ecology Letters, 6 (2003): 936-943. |
| [12] | [ M. Fan,Q. Wang,X. Zou, Dynamics of a non-autonomous ratio-dependent predator-prey system, Proceedings of the Royal Society of Edinburgh: Section A Mathematics, 133 (2003): 97-118. |
| [13] | [ P. J. S. Franks, NPZ models of plankton dynamics: Their construction, coupling to physics, and application, J. Oceanog, 58 (2002): 379-387. |
| [14] | [ J. A. Freund,S. Mieruch,B. Scholze,K. Wiltshire,U. Feudel, Bloom dynamics in a seasonally forced phytoplankton zooooplankton model: trigger mechanisms and timing effects, Ecol. Complex, 3 (2006): 129-139. |
| [15] | [ G. F. Fussmann,S. P. Ellner,K. W. Shertzer,N. G. Hairston, Crossing the hopf bifurcation in a live predator-prey system, Science, 290 (2000): 1358-1360. |
| [16] | [ R. E. Gaines and J. L. Mawhin, Coincidence Degree and Nonlinear Differential Equations, Springer-Verlag, Berlin, 1977. |
| [17] | [ R. J. Geider,H. L. Maclntyre,T. M. Kana, A dynamic regulatory model of phytoplanktonic acclimation to light, nutrients, and temperature, Limnol. Oceanogr, 43 (1998): 679-694. |
| [18] | [ P. M. Glibert, Eutrophication and Harmful Algal Blooms: A Complex Global Issue, Examples from the Arabian Seas including Kuwait Bay, and an Introduction to the Global Ecology and Oceanography of Harmful Algal Blooms (GEOHAB) Programme, Int. J. Oceans and Oceanography, 2 (2007): 157-169. |
| [19] | [ J. C. Goldman,J. C. Edward, A kinetic approach to the effect of temperature on algal growth, Limnol. Oceanogr., 19 (1974): 756-766. |
| [20] | [ G. M. Grimaud,V. L. Guennec,S. D. Ayata,F. Mariret,A. Schiandra,O. Bernard, Modelling the effect of temperature on phytoplankton growth across the global ocean, IFAC-PapersOnLine, 48 (2015): 228-233. |
| [21] | [ J. P. Grover,T. H. Chrzanowski, Seasonal dynamics of phytoplankton in two warm temperate reservoirs: association of taxonomic composition with temperature, J. Plankt. Res, 28 (2006): 1-17. |
| [22] | [ C. S. Holling, The functional response of predators to prey density and its role in mimicry and population regulation, Memoirs of the Entomological Society of Canada, 97 (1965): 5-60. |
| [23] | [ A. Huppert,B. Blasius,R. Olinky,L. Stone, A model for seasonal phytoplankton blooms, J. Theor. Biol., 236 (2005): 276-290. |
| [24] | [ K. S. Johnson,F. P. Chavez,G. E. Friederich, Continental-shelf sediment as a primary source of iron for coastal phytoplankton, Nature, 398 (1999): 697-700. |
| [25] | [ R. I. Jones, The importance of temperature conditioning to the respiration of natural phytoplankton communities, British Phycological Journal, 12 (2007): 277-285. |
| [26] | [ S. E. Jorgensen and G. Bendoricchio, Fundamentals of Ecological Modelling, Elsevier, 2001. |
| [27] | [ I. Loladze,Y. Kuang,J. J. Elser, Stoichiometry in producer-grazer systems: Linking energy flow with element cycling, Bulletin of Mathematical Biology, 62 (2000): 1137-1162. |
| [28] | [ J. H. Luo, Phytoplankton-zooplankton dynamics in periodic environments taking into account eutrophication, Math. Biosci., 245 (2013): 126-136. |
| [29] | [ J. R. Moisan,T. A. Moisan,M. R. Abbott, Modelling the effect of temperature on the maximum growth rates of phytoplankton populations, Ecol. Model., 153 (2002): 197-215. |
| [30] | [ G. Phillips,R. Jackson,C. Bennett,A. Chilvers, The importance of sediment phosphorus release in the restoration of very shallow lakes (The Norfolk Broads, England) and implications for biomanipulation, Hydrobiologia, 94 (1994): 445-456. |
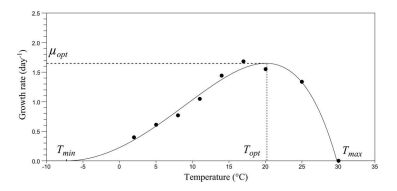
| [31] | [ L. Rosso,J. R. Lobry,J. P. Flandrois, An unexpected correlation between cardinal temperatures of microbial growth highlighted by a new model, J. Theor. Biol., 162 (1993): 447-463. |
| [32] | [ J. B. Shukla,A. K. Misra,P. Chandra, Modeling and analysis of the algal bloom in a lake caused by discharge of nutrients, App. Math. Comput., 196 (2008): 782-790. |
| [33] | [ S. J. Thackeray,I. D. Jones,S. C. Maberly, Long-term change in the phenology of spring phytoplankton: species-specific responses to nutrient enrichement and climatic changes, J. Ecol., 96 (2008): 523-535. |
| [34] | [ D. Toro,M. Dominic,D. J. O'Connor,R. V. Thomann, A dynamic model of the phytoplankton population in the Sacramento San Joaquin Delta, Adv. Chem. Ser, 106 (1971): 131-180. |
| [35] | [ C. L. Wang,W. Y. Pan,Y. Q. Han,X. Qian, Effect of global climate change on cyanobacteria bloom in taihu lake, China Environmental Science, 30 (2010): 822-828. |
| [36] | [ F. B. Wang,S. B. Hsu,W. D. Wang, Dynamics of harmful algae with seasonal temperature variations in the cove-main lake, Discrete and Continuous Dynams. Systems -B, 21 (2016): 313-315. |
| [37] | [ W. D. Wang,X. Q. Zhao, Threshold dynamics for compartmental epidemic models in periodic environments, J. Dyn. Differ. Equ., 20 (2008): 699-717. |
| [38] | [ M. Winder,D. E. Schindler, Climate change uncouples trophic interactions in an aquatic ecosystem, Ecology, 85 (2004): 2100-2106. |
| [39] | [ M. Winder,J. E. Cloern, The annual cycles of phytoplankton biomass, Philos. T. R. Soc. B, 365 (2010): 3215-3226. |
| [40] | [ Q. J. Xu,B. Q. Qin,W. M. Chen,Y. W. Chen,G. Gao, Ecological simulation of algae growth in Taihu Lake, J. Lake Sci., 2 (2001): 149-157. |
| [41] | [ F. Zhang,X. Q. Zhao, A periodic epidemic model in a patchy environment, J. Math. Anal. Appl., 325 (2007): 496-516. |
| [42] | [ Q. Zhao,X. Lu, Parameter estimation in a three-dimensional marine ecosystem model using the adjoint technique, J. Marine Syst., 74 (2008): 443-452. |
| [43] | [ X. Q. Zhao, Dynamical Systems in Population Biology, Springer-Verlag, New York, 2003. |
Figures(13) / Tables(3)

Ming Chen, Meng Fan, Xing Yuan, Huaiping Zhu. Effect of seasonal changing temperature on the growth of phytoplankton[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1091-1117. doi: 10.3934/mbe.2017057







 DownLoad:
DownLoad: