Citation: Kelum Gajamannage, Erik M. Bollt. Detecting phase transitions in collective behavior using manifold's curvature[J]. Mathematical Biosciences and Engineering, 2017, 14(2): 437-453. doi: 10.3934/mbe.2017027

| [1] | [ Birds flying away, shutterstock. Available from: https://www.shutterstock.com/video/clip-3003274-stock-footage-birds-flying-away.html?src=search/Yg-XYej1Po2F0VO3yykclw:1:19/gg. |
| [2] | [ Data set of detection of unusual crowd activity available at robotics and vision laboratory, Department of Computer Science and Engineering, University of Minnesota. Available from: http://mha.cs.umn.edu/proj_events.shtml. |
| [3] | [ Data set of pet2009 at Computational Vision Group, University of Reading, 2009. Available from: http://ftp.pets.reading.ac.uk/pub/. |
| [4] | [ N. Abaid,E. Bollt,M. Porfiri, Topological analysis of complexity in multiagent systems, Physical Review E, 85 (2012): 041907. |
| [5] | [ G. Alfred, Modern Differential Geometry of Curves and Surfaces with Mathematica, CRC press, 1998. |
| [6] | [ I. R. de Almeida and C. R. Jung, Change detection in human crowds, in Graphics, Patterns and Images (SIBGRAPI), 2013 26th SIBGRAPI-Conference on, IEEE, (2013), 63-69. |
| [7] | [ E. L. Andrade, S. Blunsden and R. B. Fisher, Hidden markov models for optical flow analysis in crowds, in Pattern Recognition, 2006. ICPR 2006. 18th International Conference on, IEEE, 1(2006), 460-463. |
| [8] | [ M. Ballerini,N. Cabibbo,R. Candelier,A. Cavagna,E. Cisbani,I. Giardina,A. Orlandi,G. Parisi,A. Procaccini,M. Viale, Empirical investigation of starling flocks: A benchmark study in collective animal behavior, Animal Behaviour, 76 (2008): 201-215. |
| [9] | [ C. Becco,N. Vandewalle,J. Delcourt,P. Poncin, Experimental evidences of a structural and dynamical transition in fish school, Physica A: Statistical Mechanics and its Applications, 367 (2006): 487-493. |
| [10] | [ M. Beekman,D. J. T. Sumpter,F. L. W. Ratnieks, Phase transition between disordered and ordered foraging in pharaoh's ants, Proceedings of the National Academy of Sciences, 98 (2001): 9703-9706. |
| [11] | [ A. C. Bovik, Handbook of Image and Video Processing, Academic press, 2010. |
| [12] | [ R. Bracewell, Fourier Analysis and Imaging, Springer Science & Business Media, 2010. |
| [13] | [ I. D. Couzin, Collective cognition in animal groups, Trends in cognitive sciences, 13 (2009): 36-43. |
| [14] | [ I. D. Couzin,J. Krause,N. R. Franks,S. A. Levin, Effective leadership and decision-making in animal groups on the move, Nature, 433 (2005): 513-516. |
| [15] | [ A. Deutsch, Principles of biological pattern formation: Swarming and aggregation viewed as self organization phenomena, Journal of Biosciences, 24 (1999): 115-120. |
| [16] | [ J. H. Friedman,J. L. Bentley,R. A. Finkel, An algorithm for finding best matches in logarithmic expected time, ACM Transactions on Mathematical Software, 3 (1977): 209-226. |
| [17] | [ K. Gajamannage,S. Butailb,M. Porfirib,E. M. Bollt, Model reduction of collective motion by principal manifolds, Physica D: Nonlinear Phenomena, 291 (2015): 62-73. |
| [18] | [ K. Gajamannage,S. Butailb,M. Porfirib,E. M. Bollt, Identifying manifolds underlying group motion in Vicsek agents, The European Physical Journal Special Topics, 224 (2015): 3245-3256. |
| [19] | [ J. J. Gerbrands, On the relationships between SVD, KLT and PCA, Pattern recognition, 14 (1981): 375-381. |
| [20] | [ R. Gerlai, High-throughput behavioral screens: The first step towards finding genes involved in vertebrate brain function using zebra fish, Molecules, 15 (2010): 2609-2622. |
| [21] | [ G. H. Golub,C. Reinsch, Singular value decomposition and least squares solutions, Numerische Mathematik, 14 (1970): 403-420. |
| [22] | [ D. Helbing,J. Keltsch,P. Molnar, Modelling the evolution of human trail systems, Nature, 388 (1997): 47-50. |
| [23] | [ J. M. Lee, Riemannian Manifolds: An Introduction to Curvature, volume 176, Springer, 1997. |
| [24] | [ J. M. Lee, Introduction to Smooth Manifolds, Graduate Texts in Mathematics, 218. Springer-Verlag, New York, 2003. |
| [25] | [ R. Mehran, A. Oyama and M. Shah, Abnormal crowd behavior detection using social force model, in Computer Vision and Pattern Recognition, 2009. CVPR 2009. IEEE Conference on, IEEE, (2009), 935-942. |
| [26] | [ M. M. Millonas, Swarms, Phase Transitions, and Collective Intelligence, Technical report, Los Alamos National Lab., New Mexico, USA, 1992. |
| [27] | [ S. R. Musse and D. Thalmann, A model of human crowd behavior: Group inter-relationship and collision detection analysis, in Computer Animation and Simulation, Springer, (1997), 39-51. |
| [28] | [ M. Nagy,Z. Ákos,D. Biro,T. Vicsek, Hierarchical group dynamics in pigeon flocks, Nature, 464 (2010): 890-893. |
| [29] | [ B. O'neill, null, Elementary Differential Geometry, Academic press, New York, 1966. |
| [30] | [ T. Papenbrock and T. H. Seligman, Invariant manifolds and collective motion in many-body systems, AIP Conf. Proc. , 597(2001), p301, arXiv: nlin/0206035. |
| [31] | [ B. L. Partridge, The structure and function of fish schools, Scientific American, 246 (1982): 114-123. |
| [32] | [ W. Rappel,A. Nicol,A. Sarkissian,H. Levine,W. F. Loomis, Self-organized vortex state in two-dimensional dictyostelium dynamics, Physical Review Letters, 83 (1999): p1247. |
| [33] | [ E. M. Rauch,M. M. Millonas,D. R. Chialvo, Pattern formation and functionality in swarm models, Physics Letters A, 207 (1995): 185-193. |
| [34] | [ V. Y. Rovenskii, Topics in Extrinsic Geometry of Codimension-one Foliations, Springer, 2011. |
| [35] | [ S. T. Roweis,L. K. Saul, Nonlinear dimensionality reduction by locally linear embedding, Science, 290 (2000): 2323-2326. |
| [36] | [ R. V. Solé,S. C. Manrubia,B. Luque,J. Delgado,J. Bascompte, Phase transitions and complex systems: Simple, nonlinear models capture complex systems at the edge of chaos, Complexity, 1 (1996): 13-26. |
| [37] | [ D. Somasundaram, Differential Geometry: A First Course, Alpha Science Int'l Ltd., 2005. |
| [38] | [ D. Sumpter,J. Buhl,D. Biro,I. Couzin, Information transfer in moving animal groups, Theory in Biosciences, 127 (2008): 177-186. |
| [39] | [ J. B. Tenenbaum,V. De Silva,J. C. Langford, A global geometric framework for nonlinear dimensionality reduction, Science, 290 (2000): 2319-2323. |
| [40] | [ E. Toffin,D. D. Paolo,A. Campo,C. Detrain,J. Deneubourg, Shape transition during nest digging in ants, Proceedings of the National Academy of Sciences, 106 (2009): 18616-18620. |
| [41] | [ C. M. Topaz,A. L. Bertozzi, Swarming patterns in a two-dimensional kinematic model for biological groups, SIAM Journal on Applied Mathematics, 65 (2004): 152-174. |
| [42] | [ T. Vicsek,A. Cziró,E. Ben-Jacob,I. Cohen,O. Shochet, Novel type of phase transition in a system of self-driven particles, Physical Review Letters, 75 (1995): 1226-1229. |
| [43] | [ E. Witten, Phase transitions in m-theory and f-theory, Nuclear Physics B, 471 (1996): 195-216. |
| [44] | [ P. N. Yianilos, Data structures and algorithms for nearest neighbor search in general metric spaces, in Proceedings of the Fourth Annual ACM-SIAM Symposium on Discrete Algorithms, Society for Industrial and Applied Mathematics, (1993), 311-321. |
| [45] | [ T. Zhao and R. Nevatia, Tracking multiple humans in crowded environment, in Computer Vision and Pattern Recognition, 2004. CVPR 2004. Proceedings of the 2004 IEEE Computer Society Conference on, IEEE, (2004), Ⅱ-406. |
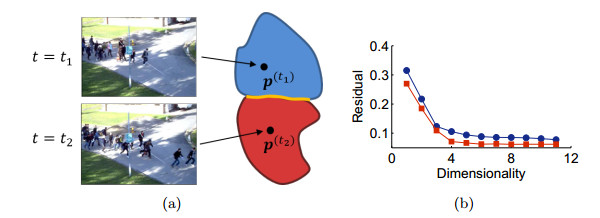
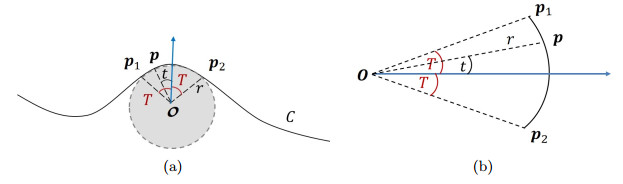
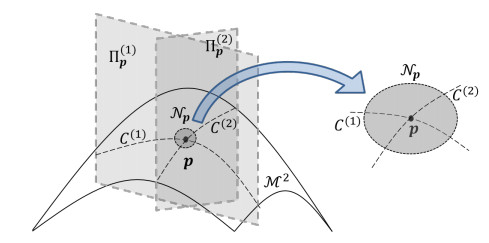
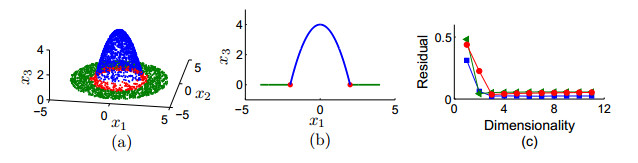
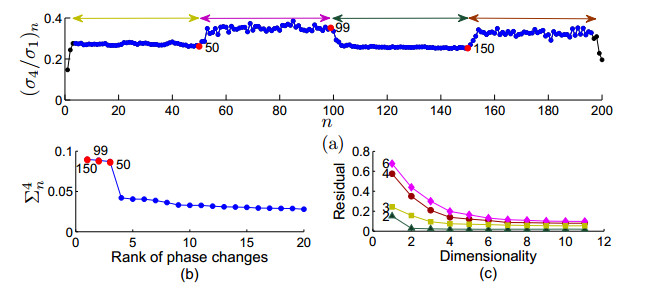
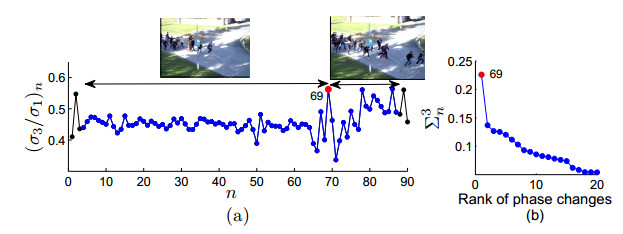
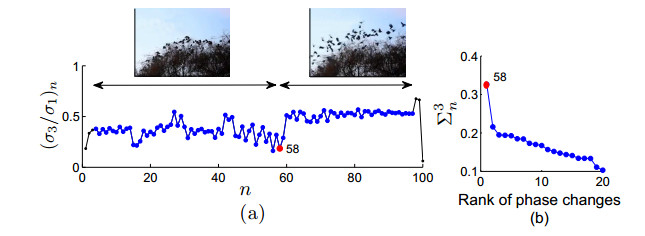
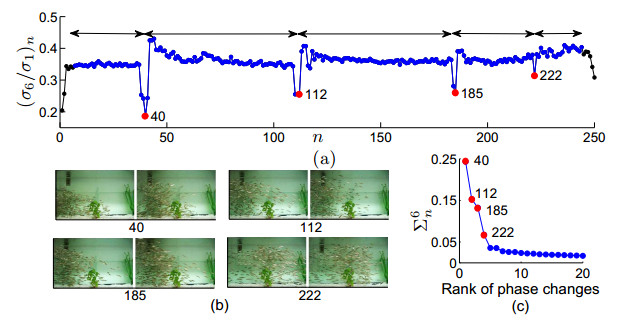
Figures(9)

Kelum Gajamannage, Erik M. Bollt. Detecting phase transitions in collective behavior using manifold's curvature[J]. Mathematical Biosciences and Engineering, 2017, 14(2): 437-453. doi: 10.3934/mbe.2017027







 DownLoad:
DownLoad: