Citation: Minette Herrera, Aaron Miller, Joel Nishimura. Altruistic aging: The evolutionary dynamics balancing longevity and evolvability[J]. Mathematical Biosciences and Engineering, 2017, 14(2): 455-465. doi: 10.3934/mbe.2017028

| [1] | [ P. A. Abrams, Does increased mortality favor the evolution of more rapid senescence?, Evolution, 47 (1993): 877-887. |
| [2] | [ P. Bak,K. Sneppen, Punctuated equilibrium and criticality in a simple model of evolution, Physical Review Letters, 71 (1993): 4083. |
| [3] | [ S. Balshine-Earn,F. C. Neat,H. Reid,M. Taborsky, Paying to stay or paying to breed? field evidence for direct benefits of helping behavior in a cooperatively breeding fish, Behavioral Ecology, 9 (1998): 432-438. |
| [4] | [ G. Bavestrello,C. Sommer,M. Sarà, Bi-directional conversion in turritopsis nutricula (hydrozoa), Scientia Marina, 56 (1992): 137-140. |
| [5] | [ J. Bendor,P. Swistak, Types of evolutionary stability and the problem of cooperation, Proceedings of the National Academy of Sciences, 92 (1995): 3596-3600. |
| [6] | [ J. D. Congdon,R. D. Nagle,O. M. Kinney,R. C. van Loben Sels,T. Quinter,D. W. Tinkle, Testing hypotheses of aging in long-lived painted turtles (chrysemys picta), Experimental Gerontology, 38 (2003): 765-772. |
| [7] | [ F. Fu,M. A. Nowak, Global migration can lead to stronger spatial selection than local migration, Journal of Statistical Physics, 151 (2013): 637-653. |
| [8] | [ L. Hadany,T. Beker,I. Eshel,M. W. Feldman, Why is stress so deadly? an evolutionary perspective, Proceedings of the Royal Society of London B: Biological Sciences, 273 (2006): 881-885. |
| [9] | [ W. Hamilton, The genetical evolution of social behaviour. Ⅰ, Journal of Theoretical Biology, 7 (1964): 1-16. |
| [10] | [ G. Ichinose,M. Saito,H. Sayama,D.S. Wilson, Adaptive long-range migration promotes cooperation under tempting conditions, Scientific Reports, 3 (2013): p2509. |
| [11] | [ K. Jin, Modern biological theories of aging, Aging and Disease, 1 (2010): p72. |
| [12] | [ M. Kimura,G.H. Weiss, The stepping stone model of population structure and the decrease of genetic correlation with distance, Genetics, 49 (1964): p561. |
| [13] | [ T. B. Kirkwood, Evolution of ageing, Mechanisms of Ageing and Development, 123 (2002): 737-745. |
| [14] | [ P. Ljubuncic,A. Z. Reznick, The evolutionary theories of aging revisited-a mini-review, Gerontology, 55 (2009): 205-216. |
| [15] | [ P. B. Medawar, An Unsolved Problem of Biology, College, 1952. |
| [16] | [ M. A. Nowak, Five rules for the evolution of cooperation, Science, 314 (2006): 1560-1563. |
| [17] | [ H. Ohtsuki,P. Bordalo,M. A. Nowak, The one-third law of evolutionary dynamics, Journal of Theoretical Biology, 249 (2007): 289-295. |
| [18] | [ L. Partridge,N. H. Barton, Optimality, mutation and the evolution of ageing, Nature: International Weekly Journal of Science, 362 (1993): 305-311. |
| [19] | [ G. B. Pollock,A. Cabrales, Suicidal altruism under random assortment, Evolutionary Ecology Research, 10 (2008): 1077-1086. |
| [20] | [ A. Traulsen,M. A. Nowak, Evolution of cooperation by multilevel selection, Proceedings of the National Academy of Sciences, 103 (2006): 10952-10955. |
| [21] | [ J. Van Cleve, Social evolution and genetic interactions in the short and long term, Theoretical Population Biology, 103 (2015): 2-26. |
| [22] | [ J. W. Vaupel, A. Baudisch, M. Dölling, D. A. Roach and J. Gampe, The case for negative senescence, Theoretical Population Biology, 65 (2004), 339-351, http://www.sciencedirect.com/science/article/pii/S004058090400022X, Demography in the 21st Century. |
| [23] | [ A. Weismann, E. B. Poulton, S. Schönland and A. E. Shipley, Essays Upon Heredity and Kindred Biological Problems, vol. 1, Clarendon press, 1891. |
| [24] | [ G. C. Williams, Pleiotropy, natural selection, and the evolution of senescence, Evolution, 11 (1957): 398-411. |
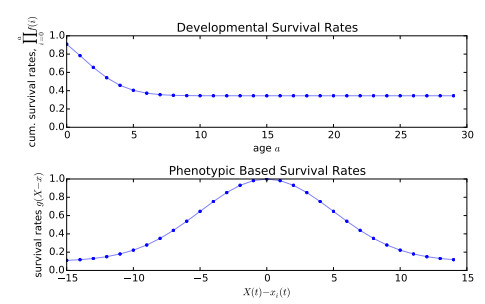
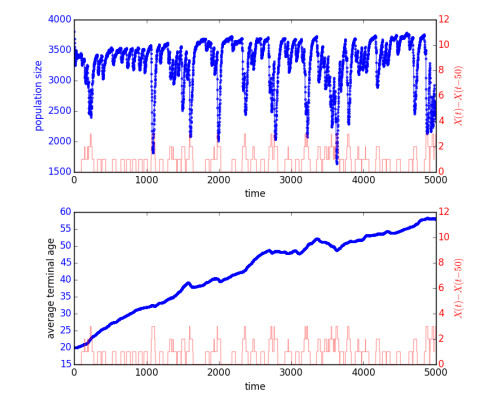
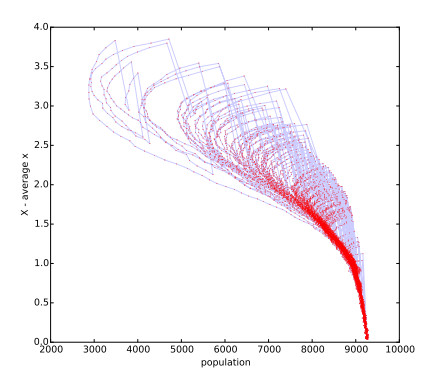
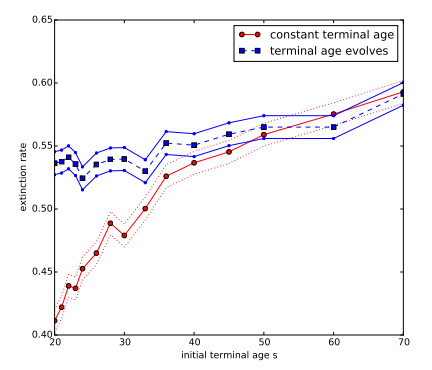
Figures(7)

Minette Herrera, Aaron Miller, Joel Nishimura. Altruistic aging: The evolutionary dynamics balancing longevity and evolvability[J]. Mathematical Biosciences and Engineering, 2017, 14(2): 455-465. doi: 10.3934/mbe.2017028







 DownLoad:
DownLoad: