Citation: Kazeem Oare Okosun, Robert Smith?. Optimal control analysis of malaria-schistosomiasis co-infection dynamics[J]. Mathematical Biosciences and Engineering, 2017, 14(2): 377-405. doi: 10.3934/mbe.2017024

| [1] | [ B. M. Adams,H. T. Banks,H. Kwon,H. T. Tran, Dynamic multidrug therapies for HIV: Optimal and STI control approaches, Mathematical Biosciences and Engineering, 1 (2004): 223-241. |
| [2] | [ F. B. Agusto, Optimal chemoprophylaxis and treatment control strategies of a tuberculosis transmission model, World Journal of Modelling and Simulation, 5 (2009): 163-173. |
| [3] | [ F. B. Agusto,K. O. Okosun, Optimal seasonal biocontrol for Eichhornia crassipes, International Journal of Biomathematics, 3 (2010): 383-397. |
| [4] | [ R. M. Anderson and R. M. May, Infectious Diseases of Humans: Dynamics and Control, Oxford University Press, 1991, Oxford. |
| [5] | [ K. W. Blayneh,Y. Cao,H. D. Kwon, Optimal control of vector-borne diseases: Treatment and Prevention, Discrete and Continuous Dynamical Systems Series B, 11 (2009): 587-611. |
| [6] | [ J. G. Breman,M. S. Alilio,A. Mills, Conquering the intolerable burden of malaria: What's new, what's needed: A summary, Am. J. Trop. Med. Hyg., 71 (2004): 1-15. |
| [7] | [ C. Castillo-Chavez,B. Song, Dynamical model of tuberculosis and their applications, Math. Biosci. Eng., 1 (2004): 361-404. |
| [8] | [ Z. Chen,L. Zou,D. Shen,W. Zhang,S. Ruan, Mathematical modelling and control of Schistosomiasis in Hubei Province, China, Acta Tropica, 115 (2010): 119-125. |
| [9] | [ E. T. Chiyaka,G. Magombedze,L. Mutimbu, Modelling within host parasite dynamics of schistosomiasis, Comp. Math. Meth. Med., 11 (2010): 255-280. |
| [10] | [ J. A. Clennon,C. G. King,E. M. Muchiri,U. Kitron, Hydrological modelling of snail dispersal patterns in Msambweni, Kenya and potential resurgence of Schistosoma haematobium transmission, Parasitology, 134 (2007): 683-693. |
| [11] | [ S. Doumbo,T. M. Tran,J. Sangala,S. Li,D. Doumtabe, Co-infection of long-term carriers of Plasmodium falciparum with Schistosoma haematobium enhances protection from febrile malaria: A prospective cohort study in Mali, PLoS Negl. Trop. Dis., 8 (2014): e3154. |
| [12] | [ M. Finkel, Malaria: Stopping a Global Killer, National Geographic, July 2007. |
| [13] | [ Z. Feng,A. Eppert,F. A. Milner,D. J. Minchella, Estimation of parameters governing the transmission dynamics of schistosomes, Appl. Math. Lett., 17 (2004): 1105-1112. |
| [14] | [ W. H. Fleming,R. W. Rishel, null, Deterministic and Stochastic Optimal Control, Springer Verlag, New York, 1975. |
| [15] | [ J. H. Ge,S. Q. Zhang,T. P. Wang,G. Zhang,C. Tao,D. Lu,Q. Wang,W. Wu, Effects of flood on the prevalence of schistosomiasis in Anhui province in 1998, Journal of Tropical Diseases and Parasitology, 2 (2004): 131-134. |
| [16] | [ P. J. Hotez,D. H. Molyneux,A. Fenwick,E. Ottesen, Ehrlich and S. Sachs et al., Incorporating a rapid-impact package for neglected tropical diseases with programs for HIV/AIDS, tuberculosis, and malaria, PLoS Med., 3 (2006): e102. |
| [17] | [ M. Y. Hyun, Comparison between schistosomiasis transmission modelings considering acquired immunity and age-structured contact pattern with infested water, Mathematical Biosciences, 184 (2003): 1-26. |
| [18] | [ H. R. Joshi, Optimal control of an HIV immunology model, Optimal Control Applications in Mathematics, 23 (2002): 199-213. |
| [19] | [ A. Kealey,R. J. Smith?, Neglected Tropical Diseases: Infection, modelling and control, J. Health Care for the Poor and Underserved, 21 (2010): 53-69. |
| [20] | [ J. Keiser,J. Utzinger,M. Caldas de Castro,T. A. Smith,M. Tanner,B. Singer, Urbanization in sub-Saharan Africa and implication for malaria control, Am. J. Trop. Med. Hyg., 71 (2004): 118-127. |
| [21] | [ D. Kirschner,S. Lenhart,S. Serbin, Optimal Control of the Chemotherapy of HIV, J. Math. Biol., 35 (1997): 775-792. |
| [22] | [ J. C. Koella,R. Anita, Epidemiological models for the spread of anti-malaria resistance, Malaria Journal, 2 (2003): p3. |
| [23] | [ C. M. Kribs-Zaleta,J. X. Velasco-Hernandez, A simple vaccination model with multiple endemic states, Math. Biosci., 164 (2000): 183-201. |
| [24] | [ V. Lakshmikantham, S. Leela and A. A. Martynyuk, Stability Analysis of Nonlinear Systems, Marcel Dekker, New York and Basel, 1989. |
| [25] | [ S. Lenhart and J. T. Workman, Control Applied to Biological Models, Chapman and Hall, London, 2007. |
| [26] | [ J. Li, D. Blakeley and R. J. Smith?, The failure of $ R_0 $, Comp. Math. Meth. Med. , 2011 (2011), Article ID 527610, 17pp. |
| [27] | [ G. Li,Z. Jin, Global stability of a SEIR epidemic model with infectious force in latent, infected and immune period, Chaos, Solutions and Fractals, 25 (2005): 1177-1184. |
| [28] | [ Q. Longxing, J. Cui, T. Huang, F. Ye and L. Jiang, Mathematical model of schistosomiasis under flood in Anhui province Abstract and Applied Analysis, 2014(2014), Article ID 972189, 7pp. |
| [29] | [ A. D. Lopez,C. D. Mathers,M. Ezzati,D. T. Jamison,C. J. Murray, Global and regional burden of disease and risk factors, 2001: Systematic analysis of population health data, Lancet, 367 (2006): 1747-1757. |
| [30] | [ E. Mtisi,H. Rwezaura,J. M. Tchuenche, A mathematical analysis of malaria and Tuberculosis co-dynamics, Discrete and Continuous Dynamical Systems Series B, 12 (2009): 827-864. |
| [31] | [ Z. Mukandavire,A. B. Gumel,W. Garira,J. M. Tchuenche, Mathematical analysis of a model for HIV-Malaria co-infection, Mathematical Biosciences and Engineering, 6 (2009): 333-362. |
| [32] | [ S. Mushayabasa and C. P. Bhunu, Modeling Schistosomiasis and HIV/AIDS co-dynamics, Computational and Mathematical Methods in Medicine, 2011(2011), Article ID 846174, 15pp. |
| [33] | [ S. Mushayabasa,C. P. Bhunu, Is HIV infection associated with an increased risk for cholera? Insights from mathematical model, Biosystems, 109 (2012): 203-213. |
| [34] | [ I. S. Nikolaos,K. Dietz,D. Schenzle, Analysis of a model for the Pathogenesis of AIDS, Mathematical Biosciences, 145 (1997): 27-46. |
| [35] | [ K. O. Okosun,R. Ouifki,N. Marcus, Optimal control analysis of a malaria disease transmission model that includes treatment and vaccination with waning immunity, BioSystems, 106 (2011): 136-145. |
| [36] | [ K. O. Okosun,O. D. Makinde, Optimal control analysis of malaria in the presence of non-linear incidence rate, Appl. Comput. Math., 12 (2013): 20-32. |
| [37] | [ K. O. Okosun,O. D. Makinde, A co-infection model of malaria and cholera diseases with optimal control, Mathematical Biosciences, 258 (2014): 19-32. |
| [38] | [ L. S. Pontryagin, V. G. Boltyanskii, R. V. Gamkrelidze and E. F. Mishchenko, The Mathematical Theory of Optimal Processes, Wiley, New York, 1962. |
| [39] | [ R. Ross, The Prevention of Malaria, Murray, London, 1911. |
| [40] | [ P. Salgame,G. S. Yap,W. C. Gause, Effect of helminth-induced immunity on infections with microbial pathogens, Nature Immunology, 14 (2013): 1118-1126. |
| [41] | [ A. A. Semenya,J. S. Sullivan,J. W. Barnwell,W. E. Secor, Schistosoma mansoni Infection Impairs Antimalaria Treatment and Immune Responses of Rhesus Macaques Infected with Mosquito-Borne Plasmodium coatneyi, Infection and Immunity, 80 (2012): 3821-3827. |
| [42] | [ K. D. Silué,G. Raso,A. Yapi,P. Vounatsou,M. Tanner,E. Ńgoran,J. Utzinger, Spatially-explicit risk profiling of Plasmodium falciparum infections at a small scale: A geostatistical modelling approach, Malaria J., 7 (2008): p111. |
| [43] | [ R. J. Smith? and S. D. Hove-Musekwa, Determining effective spraying periods to control malaria via indoor residual spraying in sub-saharan Africa Journal of Applied Mathematics and Decision Sciences, 2008(2008), Article ID 745463, 19pp. |
| [44] | [ R. W. Snow,C. A. Guerra,A. M. Noor,H. Y. Myint,S. I. Hay, The global distribution of clinical episodes of Plasmodium falciparum malaria, Nature, 434 (2005): 214-217. |
| [45] | [ R. C. Spear,A. Hubbard,S. Liang,E. Seto, Disease transmission models for public health decision making: Toward an approach for designing intervention strategies for Schistosomiasis japonica, Environ. Health Perspect., 10 (2002): 907-915. |
| [46] | [ P. van den Driessche,J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002): 29-48. |
| [47] | [ R. B. Yapi,E. Hürlimann,C. A. Houngbedji,P. B. Ndri,K. D. Silué, Infection and Co-infection with Helminths and Plasmodium among School Children in Côte d'Ivoire: Results from a National Cross-Sectional Survey, PLoS Negl. Trop. Dis., 8 (2014): e2913. |
| [48] | [ X. N. Zhou,J. G. Guo,X. H. Wu, Epidemiology of schistosomiasis in the people's republic of China, 2004, Emerging Infectious Diseases, 13 (2007): 1470-1476. |
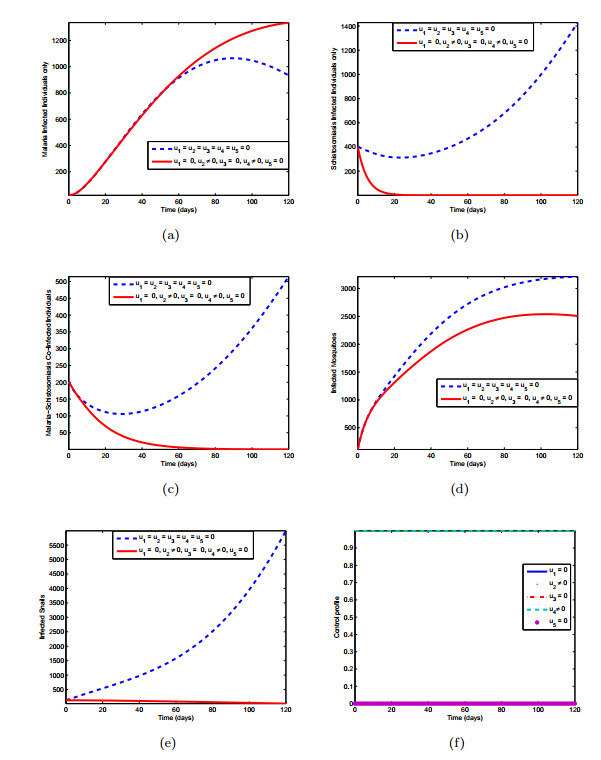
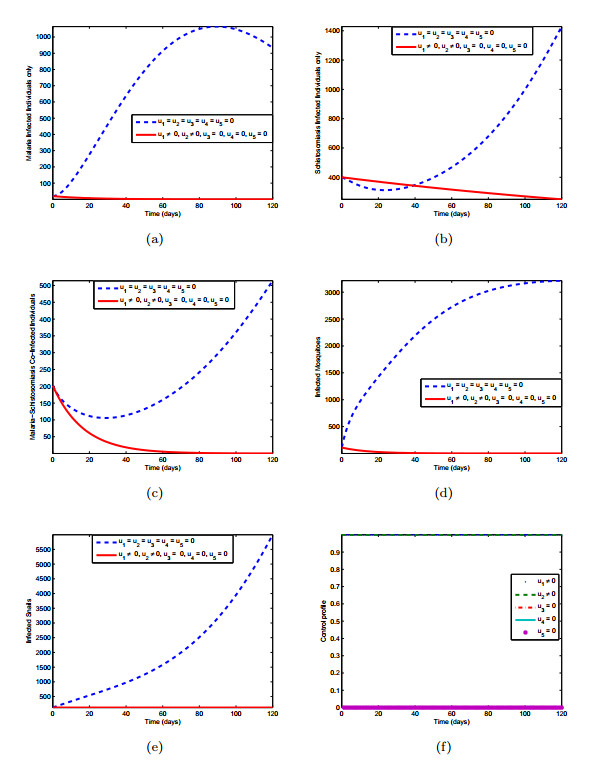
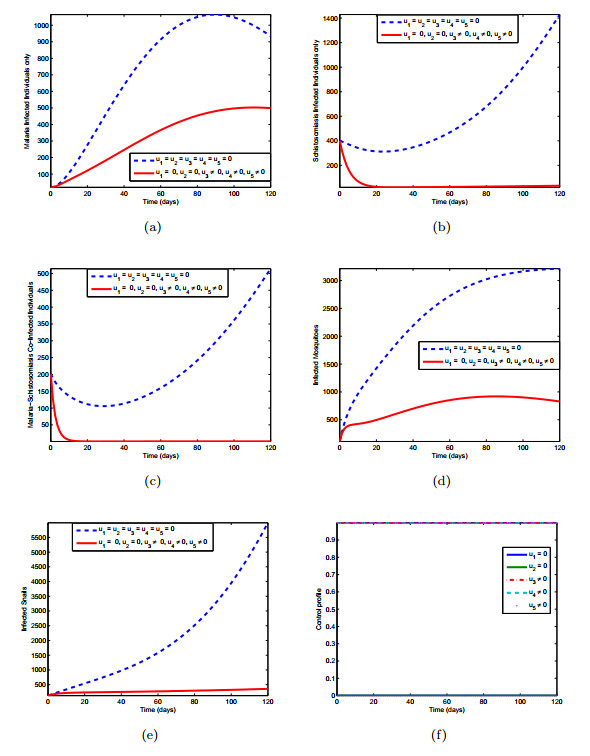
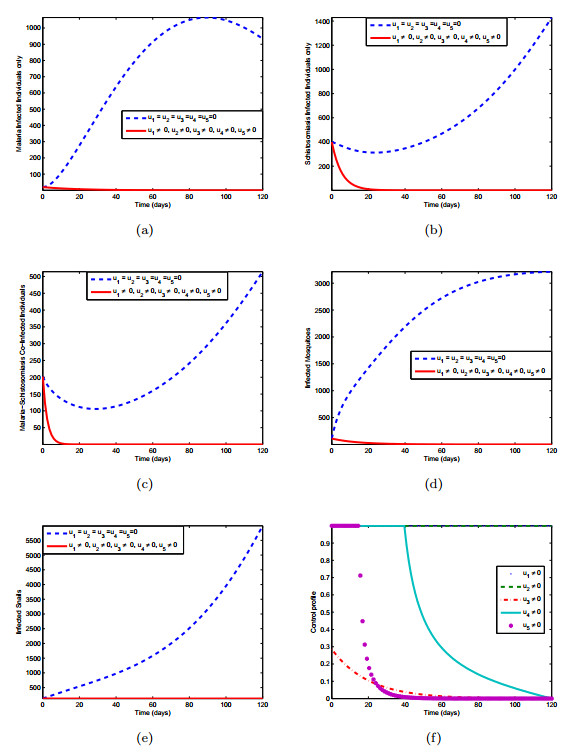
Figures(10) / Tables(3)

Kazeem Oare Okosun, Robert Smith?. Optimal control analysis of malaria-schistosomiasis co-infection dynamics[J]. Mathematical Biosciences and Engineering, 2017, 14(2): 377-405. doi: 10.3934/mbe.2017024







 DownLoad:
DownLoad: