Citation: Shuhan Jing, Adnan Younis, Dewei Chu, Sean Li. Resistive Switching Characteristics in Electrochemically Synthesized ZnO Films[J]. AIMS Materials Science, 2015, 2(2): 28-36. doi: 10.3934/matersci.2015.2.28

| [1] |
Liu SQ, Wu NJ, Ignatiev A (2000) Electric-pulse-induced reversible resistance change effect in magnetoresistive films. Appl Phys Lett 76: 2749-2751. doi: 10.1063/1.126464

|
| [2] |
Yang JJ, Pickett MD, Li X, et al. (2008) Memristive switching mechanism for metal//oxide//metal nanodevices. Nat Nano 3: 429-433. doi: 10.1038/nnano.2008.160
|
| [3] | Wu SX, Li XY, Xing XJ, et al. (2009) Resistive dependence of magnetic properties in nonvolatile Ti/Mn:TiO2/SrTi0.993Nb0.007O3/Ti memory device. Appl Phys Lett 94: 253504-253503. |
| [4] |
Beck A, Bednorz JG, Gerber C, et al. (2000) Reproducible switching effect in thin oxide films for memory applications. Appl Phys Lett 77: 139-141. doi: 10.1063/1.126902
|
| [5] |
Chua L (1971) Memristor-The missing circuit element. IEEE Transactions on Circuits Theory 18:507-519. doi: 10.1109/TCT.1971.1083337
|
| [6] |
Seo JW, Park J-W, Lim KS, et al. (2008) Transparent resistive random access memory and its characteristics for nonvolatile resistive switching. Appl Phys Lett 93: 223505-223503. doi: 10.1063/1.3041643
|
| [7] |
Strukov DB, Snider GS, Stewart DR, et al. (2008) The missing memristor found. Nature 453:80-83. doi: 10.1038/nature06932
|
| [8] |
Seo S, Lee MJ, Seo DH, et al. (2004) Reproducible resistance switching in polycrystalline NiO films. Appl Phys Lett 85: 5655-5657. doi: 10.1063/1.1831560
|
| [9] |
Choi BJ, Jeong DS, Kim SK, et al. (2005) Resistive switching mechanism of TiO2 thin films grown by atomic-layer deposition. J Appl Phys 98: 033715-033710. doi: 10.1063/1.2001146
|
| [10] |
Chen A, Haddad S, Wu YC, et al. (2008) Erasing characteristics of Cu2O metal-insulator-metal resistive switching memory. Appl Phys Lett 92: 013503-013503. doi: 10.1063/1.2828864
|
| [11] |
Villafuerte M, Heluani SP, Juarez G, et al. (2007) Electric-pulse-induced reversible resistance in doped zinc oxide thin films. Appl Phys Lett 90: 052105-052103. doi: 10.1063/1.2437688
|
| [12] |
Younis A, Chu D, Li S (2013) Bi-stable resistive switching characteristics in Ti-doped ZnO thin films. Nanoscale Res Lett 8: 1-6. doi: 10.1186/1556-276X-8-1
|
| [13] |
Sun W, Zhang K, Wang F, et al. (2013) Resistive Switching Characteristics of Zinc Oxide Resistive RAM Doped with Nickel. ECS Transactions 52: 1009-1014. doi: 10.1149/05201.1009ecst
|
| [14] |
Peng C-N, Wang C-W, Chan T-C, et al. (2012) Resistive switching of Au/ZnO/Au resistive memory: an in situ observation of conductive bridge formation. Nanoscale Res Lett 7: 559. doi: 10.1186/1556-276X-7-559
|
| [15] |
Chang W (2010) Resistive switching behaviors of ZnO nanorod layers. Appl Phys Lett 96:242109. doi: 10.1063/1.3453450
|

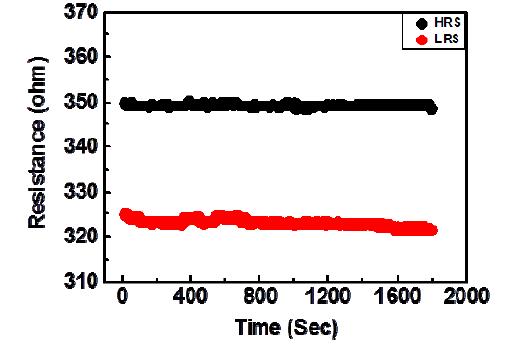
Figures(6)

Shuhan Jing, Adnan Younis, Dewei Chu, Sean Li. Resistive Switching Characteristics in Electrochemically Synthesized ZnO Films[J]. AIMS Materials Science, 2015, 2(2): 28-36. doi: 10.3934/matersci.2015.2.28







 DownLoad:
DownLoad: