Citation: Jan-Peter Hildebrandt. Pore-forming virulence factors of Staphylococcus aureus destabilize epithelial barriers-effects of alpha-toxin in the early phases of airway infection[J]. AIMS Microbiology, 2015, 1(1): 11-36. doi: 10.3934/microbiol.2015.1.11

| [1] |
Peacock SJ, de Silva I, Lowy FD (2001) What determines nasal carriage of Staphylococcus aureus? Trends Microbiol 9: 605-610. doi: 10.1016/S0966-842X(01)02254-5

|
| [2] |
Foster TJ. The Staphylococcus aureus “superbug” (2004) J Clin Invest 114: 1693-1696. doi: 10.1172/JCI200423825
|
| [3] |
Wertheim HF, Melles DC, Vos MC, et al. (2005) The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect Dis 5: 751-762. doi: 10.1016/S1473-3099(05)70295-4
|
| [4] |
Eriksen NH, Espersen F, Rosdahl VT, et al. (1995) Carriage of Staphylococcus aureus among 104 healthy persons during a 19-month period. Epidemiol Infect 115: 51-60. doi: 10.1017/S0950268800058118
|
| [5] |
Weidenmaier C, Goerke C, Wolz C (2012) Staphylococcus aureus determinants for nasal colonization. Trends Microbiol 20: 243-250. doi: 10.1016/j.tim.2012.03.004
|
| [6] |
Foster TJ (2009) Colonization and infection of the human host by staphylococci: Adhesion, survival and immune evasion. Vet Dermatol 20: 456-470. doi: 10.1111/j.1365-3164.2009.00825.x
|
| [7] |
O'Brien LM, Walsh EJ, Massey RC, et al. (2002) Staphylococcus aureus clumping factor b (Clfb) promotes adherence to human type I cytokeratin 10: Implications for nasal colonization. Cell Microbiol 4: 759-770. doi: 10.1046/j.1462-5822.2002.00231.x
|
| [8] |
Hauck CR, Ohlsen K (2006) Sticky connections: Extracellular matrix protein recognition and integrin-mediated cellular invasion by Staphylococcus aureus. Curr Opin Microbiol 9: 5-11. doi: 10.1016/j.mib.2005.12.002
|
| [9] | Mongodin E, Bajolet O, Cutrona J, et al. (2002) Fibronectin-binding proteins of Staphylococcus aureus are involved in adherence to human airway epithelium. Infect Immun 70: 620-630. |
| [10] |
Roche FM, Downer R, Keane F, et al. (2004) The N-terminal a domain of fibronectin-binding proteins A and B promotes adhesion of Staphylococcus aureus to elastin.J Biol Chem 279: 38433-38440. doi: 10.1074/jbc.M402122200
|
| [11] |
Weidenmaier C, Kokai-Kun JF, Kristian SA, et al. (2004) Role of teichoic acids in Staphylococcus aureus nasal colonization, a major risk factor in nosocomial infections. Nat Med 10: 243-245. doi: 10.1038/nm991
|
| [12] |
Patti JM, Allen BL, McGavin MJ, et al. (1994) MSCRAMM-mediated adherence of microorganisms to host tissues. Annu Rev Microbiol 48: 585-617. doi: 10.1146/annurev.mi.48.100194.003101
|
| [13] |
Clarke SR, Foster SJ (2006) Surface adhesins of Staphylococcus aureus. Adv Microb Physiol 51: 187-224. doi: 10.1016/S0065-2911(06)51004-5
|
| [14] |
van Belkum A, Melles DC, Nouwen J, et al. (2009) Co-evolutionary aspects of human colonisation and infection by Staphylococcus aureus. Infect Genet Evol 9: 32-47. doi: 10.1016/j.meegid.2008.09.012
|
| [15] |
Armstrong-Esther CA (1976) Carriage patterns of Staphylococcus aureus in a healthy non-hospital population of adults and children. Ann Hum Biol 3: 221-227. doi: 10.1080/03014467600001381
|
| [16] |
Bassetti S, Dunagana DP, D'Agostino RB, et al. (2001) Nasal carriage of Staphylococcus aureus among patients receiving allergen-injection immunotherapy: Associated factors and quantitative nasal cultures. Infect Contr Hosp Epidem 22: 741-745. doi: 10.1086/501857
|
| [17] |
Miller M, Cespedes C, Bhat M, et al. (2007) Incidence and persistence of Staphylococcus aureus nasal colonization in a community sample of HIV-infected and -uninfected drug users. Clin Infect Dis 45: 343-346. doi: 10.1086/519429
|
| [18] |
von Eiff C, Becker K, Machka K, et al. (2001) Nasal carriage as a source of Staphylococcus aureus bacteremia. Study group. N Engl J Med 344: 11-16. doi: 10.1056/NEJM200101043440102
|
| [19] |
Fritz SA, Tiemann KM, Hogan PG, et al. (2013) A serologic correlate of protective immunity against community-onset Staphylococcus aureus infection. Clin Infect Dis 56: 1554-1561. doi: 10.1093/cid/cit123
|
| [20] |
Peacock SJ, Moore CE, Justice A, et al. (2002) Virulent combinations of adhesin and toxin genes in natural populations of Staphylococcus aureus. Infect Immun 70: 4987-4996. doi: 10.1128/IAI.70.9.4987-4996.2002
|
| [21] |
Melles DC, Gorkink RF, Boelens HA, et al. (2004) Natural population dynamics and expansion of pathogenic clones of Staphylococcus aureus. J Clin Invest 114: 1732-1740. doi: 10.1172/JCI200423083
|
| [22] | Kluytmans J, van Belkum A, Verbrugh H (1997) Nasal carriage of Staphylococcus aureus: Epidemiology, underlying mechanisms, and associated risks. Clin Microbiol Rev 10: 505-520. |
| [23] |
Lee MH, Arrecubieta C, Martin FJ, et al. (2010) A postinfluenza model of Staphylococcus aureus pneumonia. J Infect Dis 201: 508-515. doi: 10.1086/650204
|
| [24] |
Handler MZ, Schwartz RA (2014) Staphylococcal scalded skin syndrome: Diagnosis and management in children and adults. J Eur Acad Dermatol Venereol 28: 1418-1423. doi: 10.1111/jdv.12541
|
| [25] | Ibler KS, Kromann CB (2014) Recurrent furunculosis - challenges and management: A review. Clin Cosm Invest Dermatol 7: 59-64. |
| [26] |
Malik Z, Roscioli E, Murphy J, et al. (2015) Staphylococcus aureus impairs the airway epithelial barrier in vitro. Int Forum Allergy Rhinol 5: 551-556. doi: 10.1002/alr.21517
|
| [27] |
Barbier F, Andremont A, Wolff M, et al. (2013) Hospital-acquired pneumonia and ventilator-associated pneumonia: Recent advances in epidemiology and management. Curr Opin Pulm Med 19: 216-228. doi: 10.1097/MCP.0b013e32835f27be
|
| [28] |
Cohen J (2002) The immunopathogenesis of sepsis. Nature 420: 885-891. doi: 10.1038/nature01326
|
| [29] |
Fournier B, Philpott DJ (2005) Recognition of Staphylococcus aureus by the innate immune system. Clin Microbiol Rev 18: 521-540. doi: 10.1128/CMR.18.3.521-540.2005
|
| [30] |
Foster TJ (2005) Immune evasion by staphylococci. Nat Rev Microbiol 3: 948-958. doi: 10.1038/nrmicro1289
|
| [31] |
Enright MC, Robinson DA, Randle G, et al. (2002) The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA). Proc Nat Acad Sci USA 99: 7687-7692. doi: 10.1073/pnas.122108599
|
| [32] |
Deresinski S (2005) Methicillin-resistant Staphylococcus aureus: An evolutionary, epidemiologic, and therapeutic odyssey. Clin Infect Dis 40: 562-573. doi: 10.1086/427701
|
| [33] |
Kahl BC (2010) Impact of Staphylococcus aureus on the pathogenesis of chronic cystic fibrosis lung disease. Int J Med Microbiol 300: 514-519. doi: 10.1016/j.ijmm.2010.08.002
|
| [34] |
Ganesan S, Comstock AT, Sajjan US (2013) Barrier function of airway tract epithelium. Tiss Barriers 1: e24997. doi: 10.4161/tisb.24997
|
| [35] |
Knowles MR, Boucher RC (2002) Mucus clearance as a primary innate defense mechanism for mammalian airways. J Clin Invest 109: 571-577. doi: 10.1172/JCI0215217
|
| [36] |
Button B, Cai LH, Ehre C, et al. (2012) A periciliary brush promotes the lung health by separating the mucus layer from airway epithelia. Science 337: 937-941. doi: 10.1126/science.1223012
|
| [37] |
Schleimer RP, Kato A, Kern R, et al. (2007) Epithelium: At the interface of innate and adaptive immune responses. J Allergy Clin Immunol 120: 1279-1284. doi: 10.1016/j.jaci.2007.08.046
|
| [38] |
Kato A, Schleimer RP (2007) Beyond inflammation: Airway epithelial cells are at the interface of innate and adaptive immunity. Curr Opin Immunol 19: 711-720. doi: 10.1016/j.coi.2007.08.004
|
| [39] |
Diamond G, Legarda D, Ryan LK (2000) The innate immune response of the respiratory epithelium. Immunol Rev 173: 27-38. doi: 10.1034/j.1600-065X.2000.917304.x
|
| [40] |
Bals R, Hiemstra PS (2004) Innate immunity in the lung: How epithelial cells fight against respiratory pathogens. Eur Resp J 23: 327-333. doi: 10.1183/09031936.03.00098803
|
| [41] |
Zaas AK, Schwartz DA (2005) Innate immunity and the lung: Defense at the interface between host and environment. Trends Cardiovasc Med 15: 195-202. doi: 10.1016/j.tcm.2005.07.001
|
| [42] |
Parker D, Prince A (2011) Innate immunity in the respiratory epithelium. Am J Respir Cell Mol Biol 45: 189-201. doi: 10.1165/rcmb.2011-0011RT
|
| [43] |
Evans SE, Xu Y, Tuvim MJ, et al. (2010) Inducible innate resistance of lung epithelium to infection. Annu Rev Physiol 72: 413-435. doi: 10.1146/annurev-physiol-021909-135909
|
| [44] |
Ausubel FM (2005) Are innate immune signaling pathways in plants and animals conserved? Nat Immunol 6: 973-979. doi: 10.1038/ni1253
|
| [45] |
Sukhithasri V, Nisha N, Biswas L, et al. (2013) Innate immune recognition of microbial cell wall components and microbial strategies to evade such recognitions. Microbiol Res 168: 396-406. doi: 10.1016/j.micres.2013.02.005
|
| [46] |
Garcia-Vallejo JJ, van Kooyk Y (2009) Endogenous ligands for C-type lectin receptors: The true regulators of immune homeostasis. Immunol Rev 230: 22-37. doi: 10.1111/j.1600-065X.2009.00786.x
|
| [47] |
Kawai T, Akira S (2007) TLR signaling. Sem Immunol 19: 24-32. doi: 10.1016/j.smim.2006.12.004
|
| [48] |
Beutler B, Jiang Z, Georgel P, et al. (2006) Genetic analysis of host resistance: Toll-like receptor signaling and immunity at large. Annu Rev Immunol 24: 353-389. doi: 10.1146/annurev.immunol.24.021605.090552
|
| [49] |
Bubeck Wardenburg J, Williams WA, Missiakas D (2006) Host defenses against Staphylococcus aureus infection require recognition of bacterial lipoproteins. Proc Nat Acad Sci USA 103: 13831-13836. doi: 10.1073/pnas.0603072103
|
| [50] |
Strober W, Murray PJ, Kitani A, et al. (2006) Signalling pathways and molecular interactions of NOD1 and NOD2. Nat Rev Immunol 6: 9-20. doi: 10.1038/nri1747
|
| [51] |
Ginsburg I (2002) Role of lipoteichoic acid in infection and inflammation. Lancet Infect Dis 2: 171-179. doi: 10.1016/S1473-3099(02)00226-8
|
| [52] |
Schwandner R, Dziarski R, Wesche H, et al. (1999) Peptidoglycan- and lipoteichoic acid-induced cell activation is mediated by Toll-like receptor 2. J Biol Chem 274: 17406-17409. doi: 10.1074/jbc.274.25.17406
|
| [53] |
Beisswenger C, Coyne CB, Shchepetov M, et al. (2007) Role of p38 MAP kinase and transforming growth factor-beta signaling in transepithelial migration of invasive bacterial pathogens. J Biol Chem 282: 28700-28708. doi: 10.1074/jbc.M703576200
|
| [54] |
Soong G, Reddy B, Sokol S, et al. (2004) TLR2 is mobilized into an apical lipid raft receptor complex to signal infection in airway epithelial cells. J Clin Invest 113: 1482-1489. doi: 10.1172/JCI200420773
|
| [55] | Inohara N, Ogura Y, Fontalba A, et al. (2003) Host recognition of bacterial muramyl dipeptide mediated through NOD2. Implications for Crohn's disease. J Biol Chem 278: 5509-5512. |
| [56] |
Gomez MI, Lee A, Reddy B, et al. (2004) Staphylococcus aureus protein a induces airway epithelial inflammatory responses by activating TNFR1. Nat Med 10: 842-848. doi: 10.1038/nm1079
|
| [57] |
Gomez MI, Prince A (2008) Airway epithelial cell signaling in response to bacterial pathogens. Pediatr Pulmonol 43: 11-19. doi: 10.1002/ppul.20735
|
| [58] | Prince AS, Mizgerd JP, Wiener-Kronish J, et al. (2006) Cell signaling underlying the pathophysiology of pneumonia. Am J Physiol 291: L297-L300. |
| [59] |
Cheon IS, Woo SS, Kang SS, et al. (2008) Peptidoglycan-mediated IL-8 expression in human alveolar type II epithelial cells requires lipid raft formation and MAPK activation. Mol Immunol 45: 1665-1673. doi: 10.1016/j.molimm.2007.10.001
|
| [60] |
Liu L, Mul FP, Lutter R, et al. (1996) Transmigration of human neutrophils across airway epithelial cell monolayers is preferentially in the physiologic basolateral-to-apical direction. Am J Respir Cell Mol Biol 15: 771-780. doi: 10.1165/ajrcmb.15.6.8969272
|
| [61] |
Kishimoto T (2010) IL-6: From its discovery to clinical applications. Int Immunol 22: 347-352. doi: 10.1093/intimm/dxq030
|
| [62] |
Gauldie J, Richards C, Harnish D, et al. (1987) Interferon beta 2/B-cell stimulatory factor type 2 shares identity with monocyte-derived hepatocyte-stimulating factor and regulates the major acute phase protein response in liver cells. Proc Nat Acad Sci USA 84: 7251-7255. doi: 10.1073/pnas.84.20.7251
|
| [63] |
Ruminy P, Gangneux C, Claeyssens S, et al. (2001) Gene transcription in hepatocytes during the acute phase of a systemic inflammation: From transcription factors to target genes. Inflamm Res 50: 383-390. doi: 10.1007/PL00000260
|
| [64] |
Chavez-Galan L, Arenas-Del Angel MC, et al. (2009) Cell death mechanisms induced by cytotoxic lymphocytes. Cell Mol Immunol 6: 15-25. doi: 10.1038/cmi.2009.3
|
| [65] | Moreilhon C, Gras D, Hologne C, et al. (2005) Live Staphylococcus aureus and bacterial soluble factors induce different transcriptional responses in human airway cells. Physiol Genom 20: 244-255. |
| [66] | Below S, Konkel A, Zeeck C, et al. (2009) Virulence factors of Staphylococcus aureus induce Erk-MAP kinase activation and c-fos expression in S9 and 16HBE14o- human airway epithelial cells. Am J Physiol 296: L470-L479. |
| [67] | Räth S, Ziesemer S, Witte A, et al. (2013) S. aureus hemolysin A-induced IL-8 and IL-6 release from human airway epithelial cells is mediated by activation of p38- and Erk-MAP kinases and additional, cell-type specific signalling mechanisms. Cell Microbiol 15: 1253-1265. |
| [68] | Rose F, Dahlem G, Guthmann B, et al. (2002) Mediator generation and signaling events in alveolar epithelial cells attacked by S. aureus alpha-toxin. Am J Physiol 282: L207-L214. |
| [69] |
Bartlett AH, Foster TJ, Hayashida A, et al. (2008) Alpha-toxin facilitates the generation of CXC chemokine gradients and stimulates neutrophil homing in Staphylococcus aureus pneumonia. J Infect Dis 198: 1529-1535. doi: 10.1086/592758
|
| [70] |
Liang X, Ji Y (2007) Involvement of alpha5beta1-integrin and TNF-alpha in Staphylococcus aureus alpha-toxin-induced death of epithelial cells. Cell Microbiol 9: 1809-1821. doi: 10.1111/j.1462-5822.2007.00917.x
|
| [71] |
Ventura CL, Higdon R, Hohmann L, et al. (2008) Staphylococcus aureus elicits marked alterations in the airway proteome during early pneumonia. Infect Immun 76: 5862-5872. doi: 10.1128/IAI.00865-08
|
| [72] |
Ventura CL, Higdon R, Kolker E, et al. (2008) Host airway proteins interact with Staphylococcus aureus during early pneumonia. Infect Immun 76: 888-898. doi: 10.1128/IAI.01301-07
|
| [73] |
McElroy MC, Cain DJ, Tyrrell C, et al. (2002) Increased virulence of a fibronectin-binding protein mutant of Staphylococcus aureus in a rat model of pneumonia. Infect Immun 70: 3865-3873. doi: 10.1128/IAI.70.7.3865-3873.2002
|
| [74] |
Dziewanowska K, Carson AR, Patti JM, et al. (2000) Staphylococcal fibronectin binding protein interacts with heat shock protein 60 and integrins: Role in internalization by epithelial cells. Infect Immun 68: 6321-6328. doi: 10.1128/IAI.68.11.6321-6328.2000
|
| [75] | Surmann K, Michalik S, Hildebrandt P, et al. (2014) Comparative proteome analysis reveals conserved and specific adaptation patterns of Staphylococcus aureus after internalization by different types of human non-professional phagocytic host cells. Front Microbiol 5: 392. |
| [76] |
Sinha B, Fraunholz M (2010) Staphylococcus aureus host cell invasion and post-invasion events. Int J Med Microbiol 300: 170-175. doi: 10.1016/j.ijmm.2009.08.019
|
| [77] |
Schnaith A, Kashkar H, Leggio SA, et al. (2007) Staphylococcus aureus subvert autophagy for induction of caspase-independent host cell death. J Biol Chem 282: 2695-2706. doi: 10.1074/jbc.M609784200
|
| [78] |
Schmidt F, Scharf SS, Hildebrandt P, et al. (2010) Time-resolved quantitative proteome profiling of host-pathogen interactions: The response of Staphylococcus aureus RN1HG to internalisation by human airway epithelial cells. Proteomics 10: 2801-2811. doi: 10.1002/pmic.201000045
|
| [79] |
Kahl BC, Goulian M, van Wamel W, et al. (2000) Staphylococcus aureus RN6390 replicates and induces apoptosis in a pulmonary epithelial cell line. Infect Immun 68: 5385-5392. doi: 10.1128/IAI.68.9.5385-5392.2000
|
| [80] |
Garzoni C, Kelley WL (2009) Staphylococcus aureus: New evidence for intracellular persistence. Trends Microbiol 17: 59-65. doi: 10.1016/j.tim.2008.11.005
|
| [81] |
Tuchscherr L, Medina E, Hussain M, et al. (2011) Staphylococcus aureus phenotype switching: An effective bacterial strategy to escape host immune response and establish a chronic infection. EMBO Mol Med 3: 129-141. doi: 10.1002/emmm.201000115
|
| [82] |
Phillips JR, Tripp TJ, Regelmann WE, et al. (2006) Staphylococcal alpha-toxin causes increased tracheal epithelial permeability. Pediatr Pulmonol 41: 1146-1152. doi: 10.1002/ppul.20501
|
| [83] |
Richter E, Harms M, Ventz K, et al. (2015) A multi-omics approach identifies key hubs associated with cell type-specific responses of airway epithelial cells to staphylococcal alpha-toxin. PLoS ONE 10: e0122089. doi: 10.1371/journal.pone.0122089
|
| [84] |
Hermann I, Räth S, Ziesemer S, et al. (2015) Staphylococcus aureus-hemolysin A disrupts cell-matrix adhesions in human airway epithelial cells. Am J Respir Cell Mol Biol 52: 14-24. doi: 10.1165/rcmb.2014-0082OC
|
| [85] |
Ratner AJ, Bryan R, Weber A, et al. (2001) Cystic fibrosis pathogens activate Ca2+-dependent mitogen-activated protein kinase signaling pathways in airway epithelial cells. J Biol Chem 276: 19267-19275. doi: 10.1074/jbc.M007703200
|
| [86] | Greene CM, Ramsay H, Wells RJ, et al. (2010) Inhibition of Toll-like receptor 2-mediated interleukin-8 production in cystic fibrosis airway epithelial cells via the alpha7-nicotinic acetylcholine receptor. Mediat Inflamm 2010: 423241. |
| [87] |
Sibbald MJ, Ziebandt AK, Engelmann S, et al. (2006) Mapping the pathways to staphylococcal pathogenesis by comparative secretomics. Microbiol Mol Biol Rev 70: 755-788. doi: 10.1128/MMBR.00008-06
|
| [88] |
Kuroda M, Ohta T, Uchiyama I, et al. (2001) Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet 357: 1225-1240. doi: 10.1016/S0140-6736(00)04403-2
|
| [89] |
Gill SR, Fouts DE, Archer GL, et al. (2005) Insights on evolution of virulence and resistance from the complete genome analysis of an early methicillin-resistant Staphylococcus aureus strain and a biofilm-producing methicillin-resistant Staphylococcus epidermidis strain. J Bacteriol 187: 2426-2438. doi: 10.1128/JB.187.7.2426-2438.2005
|
| [90] |
Baba T, Takeuchi F, Kuroda M, et al. (2002) Genome and virulence determinants of high virulence community-acquired MRSA. Lancet 359: 1819-1827. doi: 10.1016/S0140-6736(02)08713-5
|
| [91] |
Diep BA, Carleton HA, Chang RF, et al. (2006) Roles of 34 virulence genes in the evolution of hospital- and community-associated strains of methicillin-resistant Staphylococcus aureus. J Infect Dis 193: 1495-1503. doi: 10.1086/503777
|
| [92] |
Goerke C, Wolz C (2010) Adaptation of Staphylococcus aureus to the cystic fibrosis lung. Int J Med Microbiol 300: 520-525. doi: 10.1016/j.ijmm.2010.08.003
|
| [93] |
Goerke C, Pantucek R, Holtfreter S, et al. (2009) Diversity of prophages in dominant Staphylococcus aureus clonal lineages. J Bacteriol 191: 3462-3468. doi: 10.1128/JB.01804-08
|
| [94] |
Novick RP (2003) Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol Microbiol 48: 1429-1249. doi: 10.1046/j.1365-2958.2003.03526.x
|
| [95] |
Cheung AL, Koomey JM, Butler CA, et al. (1992) Regulation of exoprotein expression in Staphylococcus aureus by a locus (sar) distinct from agr. Proc Nat Acad Sci USA 89: 6462-6466. doi: 10.1073/pnas.89.14.6462
|
| [96] |
Rogasch K, Rühmling V, Pane-Farre J, et al. (2006) Influence of the two-component system SaeRS on global gene expression in two different Staphylococcus aureus strains. J Bacteriol 188: 7742-7758. doi: 10.1128/JB.00555-06
|
| [97] |
Geisinger E, George EA, Muir TW, et al. (2008) Identification of ligand specificity determinants in AgrC, the Staphylococcus aureus quorum-sensing receptor. J Biol Chem 283: 8930-8938. doi: 10.1074/jbc.M710227200
|
| [98] |
Jensen RO, Winzer K, Clarke SR, et al. (2008) Differential recognition of Staphylococcus aureus quorum-sensing signals depends on both extracellular loops 1 and 2 of the transmembrane sensor AgrC. J Mol Biol 381: 300-309. doi: 10.1016/j.jmb.2008.06.018
|
| [99] |
Heyer G, Saba S, Adamo R, et al. (2002) Staphylococcus aureusagr and sarA functions are required for invasive infection but not inflammatory responses in the lung. Infect Immun 70: 127-133. doi: 10.1128/IAI.70.1.127-133.2002
|
| [100] |
Haslinger-Löffler B, Kahl BC, Grundmeier M, et al. (2005) Multiple virulence factors are required for Staphylococcus aureus-induced apoptosis in endothelial cells. Cell Microbiol 7: 1087-1097. doi: 10.1111/j.1462-5822.2005.00533.x
|
| [101] |
Jones RC, Deck J, Edmondson RD, et al. (2008) Relative quantitative comparisons of the extracellular protein profiles of Staphylococcus aureus UAMS-1 and its sarA, agr, and sarA agr regulatory mutants using one-dimensional polyacrylamide gel electrophoresis and nanocapillary liquid chromatography coupled with tandem mass spectrometry. J Bacteriol 190: 5265-5278. doi: 10.1128/JB.00383-08
|
| [102] |
Wright JS, Jin R, Novick RP (2005) Transient interference with staphylococcal quorum sensing blocks abscess formation. Proc Nat Acad Sci USA 102: 1691-1696. doi: 10.1073/pnas.0407661102
|
| [103] |
Feng Y, Chen CJ, Su LH, et al. (2008) Evolution and pathogenesis of Staphylococcus aureus: Lessons learned from genotyping and comparative genomics. FEMS Microbiol Rev 32: 23-37. doi: 10.1111/j.1574-6976.2007.00086.x
|
| [104] |
Hecker M, Becher D, Fuchs S, et al. (2010) A proteomic view of cell physiology and virulence of Staphylococcus aureus. Int J Med Microbiol 300: 76-87. doi: 10.1016/j.ijmm.2009.10.006
|
| [105] |
Becher D, Hempel K, Sievers S, et al. (2009) A proteomic view of an important human pathogen-towards the quantification of the entire Staphylococcus aureus proteome. PLoS ONE 4: e8176. doi: 10.1371/journal.pone.0008176
|
| [106] | Ziebandt A-K, Weber H, Rudolph J, et al. (2001) Extracellular proteins of Staphylococcus aureus and the role of SarA and σB. Proteomics 1: 480-493. |
| [107] |
Ziebandt A-K, Becher D, Ohlsen K, et al. (2004) The influence of agr and σB in growth phase dependent regulation of virulence factors in Staphylococcus aureus. Proteomics 4: 3034-3047. doi: 10.1002/pmic.200400937
|
| [108] |
Recsei P, Kreiswirth B, O'Reilly M, et al. (1986) Regulation of exoprotein gene expression in Staphylococcus aureus by agr. Mol Gen Genet 202: 58-61. doi: 10.1007/BF00330517
|
| [109] | Peng HL, Novick RP, Kreiswirth B, et al. (1988) Cloning, characterization, and sequencing of an accessory gene regulator (agr) in Staphylococcus aureus. J Bacteriol 170: 4365-4372. |
| [110] | Cheung AL, Projan SJ (1994) Cloning and sequencing of sarA of Staphylococcus aureus, a gene required for the expression of agr. J Bacteriol 176: 4168-4172. |
| [111] | McCarthy H, Rudkin JK, Black NS, et al. (2015) Methicillin resistance and the biofilm phenotype in Staphylococcus aureus. Front Cell Infect Microbiol 5:1. |
| [112] |
Liang X, Ji Y (2007) Comparative analysis of staphylococcal adhesion and internalization by epithelial cells. Meth Mol Biol (Clifton, NJ) 391: 145-151. doi: 10.1007/978-1-59745-468-1_11
|
| [113] |
Jin T, Bokarewa M, Foster T, et al. (2004) Staphylococcus aureus resists human defensins by production of staphylokinase, a novel bacterial evasion mechanism. J Immunol 172: 1169-1176. doi: 10.4049/jimmunol.172.2.1169
|
| [114] |
Lan L, Cheng A, Dunman PM, et al. (2010) Golden pigment production and virulence gene expression are affected by metabolisms in Staphylococcus aureus. J Bacteriol 192: 3068-3077. doi: 10.1128/JB.00928-09
|
| [115] |
Hammel M, Sfyroera G, Pyrpassopoulos S, et al. (2007) Characterization of Ehp, a secreted complement inhibitory protein from Staphylococcus aureus. J Biol Chem 282: 30051-30061. doi: 10.1074/jbc.M704247200
|
| [116] |
de Haas CJ, Veldkamp KE, Peschel A, et al. (2004) Chemotaxis inhibitory protein of Staphylococcus aureus, a bacterial antiinflammatory agent. J Exp Med 199: 687-695. doi: 10.1084/jem.20031636
|
| [117] | Rogolsky M (1979) Nonenteric toxins of Staphylococcus aureus. Microbiol Rev 43: 320-360. |
| [118] | Prevost G, Mourey L, Colin DA, et al. (2001) Staphylococcal pore-forming toxins. Curr Top Microbiol Immunol 257: 53-83. |
| [119] |
Herbert S, Ziebandt AK, Ohlsen K, et al. (2010) Repair of global regulators in Staphylococcus aureus 8325 and comparative analysis with other clinical isolates. Infect Immun 78: 2877-2889. doi: 10.1128/IAI.00088-10
|
| [120] |
Pocsfalvi G, Cacace G, Cuccurullo M, et al. (2008) Proteomic analysis of exoproteins expressed by enterotoxigenic Staphylococcus aureus strains. Proteomics 8: 2462-2476. doi: 10.1002/pmic.200700965
|
| [121] |
Dinges MM, Orwin PM, Schlievert PM (2000) Exotoxins of Staphylococcus aureus. Clin Microbiol Rev 13: 16-34. doi: 10.1128/CMR.13.1.16-34.2000
|
| [122] |
Defres S, Marwick C, Nathwani D (2009) MRSA as a cause of lung infection including airway infection, community-acquired pneumonia and hospital-acquired pneumonia. Eur Respir J 34: 1470-1476. doi: 10.1183/09031936.00122309
|
| [123] |
Watkins RR, David MZ, Salata RA (2012) Current concepts on the virulence mechanisms of methicillin-resistant Staphylococcus aureus. J Med Microbiol 61: 1179-1193. doi: 10.1099/jmm.0.043513-0
|
| [124] |
Bubeck Wardenburg J, Bae T, Otto M, et al. (2007) Poring over pores: Alpha-hemolysin and Panton-Valentine leukocidin in Staphylococcus aureus pneumonia. Nat Med 13: 1405-1406. doi: 10.1038/nm1207-1405
|
| [125] |
Parker D, Prince A (2012) Immunopathogenesis of Staphylococcus aureus pulmonary infection. Sem Immunopathol 34: 281-297. doi: 10.1007/s00281-011-0291-7
|
| [126] |
Gillet Y, Issartel B, Vanhems P, et al. (2002) Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet 359: 753-759. doi: 10.1016/S0140-6736(02)07877-7
|
| [127] |
Labandeira-Rey M, Couzon F, Boisset S, et al. (2007) Staphylococcus aureus Panton-Valentine leukocidin causes necrotizing pneumonia. Science 315: 1130-1133. doi: 10.1126/science.1137165
|
| [128] |
Ragle BE, Bubeck Wardenburg J (2009) Anti-alpha-hemolysin monoclonal antibodies mediate protection against Staphylococcus aureus pneumonia. Infect Immun 77: 2712-2718. doi: 10.1128/IAI.00115-09
|
| [129] |
Ragle BE, Karginov VA, Bubeck Wardenburg J (2010) Prevention and treatment of Staphylococcus aureus pneumonia with a beta-cyclodextrin derivative. Antimicrob Agents Chemother 54: 298-304. doi: 10.1128/AAC.00973-09
|
| [130] |
Bubeck Wardenburg J, Schneewind O (2008) Vaccine protection against Staphylococcus aureus pneumonia. J Exp Med 205: 287-294. doi: 10.1084/jem.20072208
|
| [131] | Stulik L, Malafa S, Hudcova J, et al. (2014) Α-hemolysin activity of methicillin-susceptible S. aureus predicts ventilator-associated pneumonia. Am J Respir Crit Care Med 190: 1139-1148. |
| [132] |
Löffler B, Hussain M, Grundmeier M, et al. (2010) Staphylococcus aureus Panton-Valentine leukocidin is a very potent cytotoxic factor for human neutrophils. PLoS Pathog 6: e1000715. doi: 10.1371/journal.ppat.1000715
|
| [133] |
Genestier AL, Michallet MC, Prevost G, et al. (2005) Staphylococcus aureus Panton-Valentine leukocidin directly targets mitochondria and induces Bax-independent apoptosis of human neutrophils. J Clin Invest 115: 3117-3127. doi: 10.1172/JCI22684
|
| [134] |
Melles DC, van Leeuwen WB, Boelens HA, et al. (2006) Panton-Valentine leukocidin genes in Staphylococcus aureus. Emerg Infect Dis 12: 1174-1175. doi: 10.3201/eid1207.050865
|
| [135] | da Silva MC, Zahm JM, Gras D, et al. (2004) Dynamic interaction between airway epithelial cells and Staphylococcus aureus. Am J Physiol 287: L543-L551. |
| [136] | Hildebrand A, Pohl M, Bhakdi S (1991) Staphylococcus aureus alpha-toxin. Dual mechanism of binding to target cells. J Biol Chem 266: 17195-17200. |
| [137] | Tweten RK, Christianson KK, Iandolo JJ (1983) Transport and processing of staphylococcal alpha-toxin. J Bacteriol 156: 524-528. |
| [138] | Gray GS, Kehoe M (1984) Primary sequence of the alpha-toxin gene from Staphylococcus aureus Wood 46. Infect Immun 46: 615-618. |
| [139] |
Schwiering M, Brack A, Stork R, et al. (2013) Lipid and phase specificity of alpha-toxin from S. aureus. Biochim Biophys Acta 1828: 1962-1972. doi: 10.1016/j.bbamem.2013.04.005
|
| [140] |
Valeva A, Hellmann N, Walev I, et al. (2006) Evidence that clustered phosphocholine head groups serve as sites for binding and assembly of an oligomeric protein pore. J Biol Chem 281: 26014-26021. doi: 10.1074/jbc.M601960200
|
| [141] |
Galdiero S, Gouaux E (2004) High resolution crystallographic studies of alpha-hemolysin-phospholipid complexes define heptamer-lipid head group interactions: Implication for understanding protein-lipid interactions. Prot Sci 13: 1503-1511. doi: 10.1110/ps.03561104
|
| [142] |
Wilke GA, Bubeck Wardenburg J (2010) Role of a disintegrin and metalloprotease 10 in Staphylococcus aureus alpha-hemolysin-mediated cellular injury. Proc Nat Acad Sci USA 107: 13473-13478. doi: 10.1073/pnas.1001815107
|
| [143] |
Inoshima I, Inoshima N, Wilke GA, et al. (2011) A Staphylococcus aureus pore-forming toxin subverts the activity of ADAM10 to cause lethal infection in mice. Nat Med 17: 1310-1314. doi: 10.1038/nm.2451
|
| [144] |
Pany S, Vijayvargia R, Krishnasastry MV (2004) Caveolin-1 binding motif of alpha-hemolysin: Its role in stability and pore formation. Biochem Biophys Res Commun 322: 29-36. doi: 10.1016/j.bbrc.2004.07.073
|
| [145] |
Berube BJ, Bubeck Wardenburg J (2013) Staphylococcus aureus alpha-toxin: Nearly a century of intrigue. Toxins (Basel) 5: 1140-1166. doi: 10.3390/toxins5061140
|
| [146] | Korchev YE, Alder GM, Bakhramov A, et al. (1995) Staphylococcus aureus alpha-toxin-induced pores: Channel-like behavior in lipid bilayers and patch clamped cells. J Membr Biol 143: 143-151. |
| [147] |
Krasilnikov OV, Merzlyak PG, Yuldasheva LN, et al. (2000) Electrophysiological evidence for heptameric stoichiometry of ion channels formed by Staphylococcus aureus alpha-toxin in planar lipid bilayers. Mol Microbiol 37: 1372-1378. doi: 10.1046/j.1365-2958.2000.02080.x
|
| [148] |
Gouaux JE, Braha O, Hobaugh MR, et al. (1994) Subunit stoichiometry of staphylococcal alpha-hemolysin in crystals and on membranes: A heptameric transmembrane pore. Proc Nat Acad Sci USA 91: 12828-12831. doi: 10.1073/pnas.91.26.12828
|
| [149] |
Gouaux E (1998) Alpha-hemolysin from Staphylococcus aureus: An archetype of beta-barrel, channel-forming toxins. J Struct Biol 121: 110-122. doi: 10.1006/jsbi.1998.3959
|
| [150] |
Montoya M, Gouaux E (2003) Beta-barrel membrane protein folding and structure viewed through the lens of alpha-hemolysin. Biochim Biophys Acta 1609: 19-27. doi: 10.1016/S0005-2736(02)00663-6
|
| [151] |
Jayasinghe L, Miles G, Bayley H (2006) Role of the amino latch of staphylococcal alpha-hemolysin in pore formation: A co-operative interaction between the N terminus and position 217. J Biol Chem 281: 2195-2204. doi: 10.1074/jbc.M510841200
|
| [152] |
Valeva A, Walev I, Pinkernell M, et al. (1997) Transmembrane beta-barrel of staphylococcal alpha-toxin forms in sensitive but not in resistant cells. Proc Nat Acad Sci USA 94: 11607-11611. doi: 10.1073/pnas.94.21.11607
|
| [153] | Menzies BE, Kernodle DS (1994) Site-directed mutagenesis of the alpha-toxin gene of Staphylococcus aureus: Role of histidines in toxin activity in vitro and in a murine model. Infect Immun 62: 1843-1847. |
| [154] | Bhakdi S, Tranum-Jensen J (1991) Alpha-toxin of Staphylococcus aureus. Microbiol Rev 55: 733-751. |
| [155] |
Füssle R, Bhakdi S, Sziegoleit A, et al. (1981) On the mechanism of membrane damage by Staphylococcus aureus alpha-toxin. J Cell Biol 91: 83-94. doi: 10.1083/jcb.91.1.83
|
| [156] |
Song L, Hobaugh MR, Shustak C, et al. (1996) Structure of staphylococcal alpha-hemolysin, a heptameric transmembrane pore. Science 274: 1859-1866. doi: 10.1126/science.274.5294.1859
|
| [157] |
Menestrina G (1986) Ionic channels formed by Staphylococcus aureus alpha-toxin: Voltage-dependent inhibition by divalent and trivalent cations. J Membr Biol 90: 177-190. doi: 10.1007/BF01869935
|
| [158] |
Kasianowicz JJ, Brandin E, Branton D, et al. (1996) Characterization of individual polynucleotide molecules using a membrane channel. Proc Nat Acad Sci USA 93: 13770-13773. doi: 10.1073/pnas.93.24.13770
|
| [159] |
Aksimentiev A, Schulten K (2005) Imaging alpha-hemolysin with molecular dynamics: Ionic conductance, osmotic permeability, and the electrostatic potential map. Biophys J 88: 3745-3761. doi: 10.1529/biophysj.104.058727
|
| [160] | Walev I, Martin E, Jonas D, et al. (1993) Staphylococcal alpha-toxin kills human keratinocytes by permeabilizing the plasma membrane for monovalent ions. Infect Immun 61: 4972-4979. |
| [161] |
Kloft N, Busch T, Neukirch C, et al. (2009) Pore-forming toxins activate MAPK p38 by causing loss of cellular potassium. Biochem Biophys Res Commun 385: 503-506. doi: 10.1016/j.bbrc.2009.05.121
|
| [162] | Jonas D, Walev I, Berger T, et al. (1994) Novel path to apoptosis: Small transmembrane pores created by staphylococcal alpha-toxin in T lymphocytes evoke internucleosomal DNA degradation. Infect Immun 62: 1304-1312. |
| [163] |
Valeva A, Walev I, Gerber A, et al. (2000) Staphylococcal alpha-toxin: Repair of a calcium-impermeable pore in the target cell membrane. Mol Microbiol 36: 467-476. doi: 10.1046/j.1365-2958.2000.01865.x
|
| [164] |
Walev I, Palmer M, Martin E, et al. (1994) Recovery of human fibroblasts from attack by the pore-forming alpha-toxin of Staphylococcus aureus. Microb Pathogen 17: 187-201. doi: 10.1006/mpat.1994.1065
|
| [165] | Ahnert-Hilger G, Bhakdi S, Gratzl M (1985) Minimal requirements for exocytosis. A study using PC 12 cells permeabilized with staphylococcal alpha-toxin. J Biol Chem 260: 12730-12734. |
| [166] | Suttorp N, Seeger W, Dewein E, et al. (1985) Staphylococcal alpha-toxin-induced PGI2 production in endothelial cells: Role of calcium. Am J Physiol 248: C127-C134. |
| [167] |
Eichstaedt S, Gäbler K, Below S, et al. (2009) Effects of Staphylococcus aureus-hemolysin a on calcium signalling in immortalized human airway epithelial cells. Cell Calcium 45: 165-176. doi: 10.1016/j.ceca.2008.09.001
|
| [168] |
Gierok P, Harms M, Richter E, et al. (2014) Staphylococcus aureus alpha-toxin mediates general and cell type-specific changes in metabolite concentrations of immortalized human airway epithelial cells. PLoS ONE 9: e94818. doi: 10.1371/journal.pone.0094818
|
| [169] | Husmann M, Dersch K, Bobkiewicz W, et al. (2006) Differential role of p38 mitogen activated protein kinase for cellular recovery from attack by pore-forming S. aureus alpha-toxin or streptolysin O. Biochem Biophys Res Commun 344: 1128-1134. |
| [170] | Ostedgaard LS, Shasby DM, Welsh MJ (1992) Staphylococcus aureus alpha-toxin permeabilizes the basolateral membrane of a Cl--secreting epithelium. Am J Physiol 263: L104-L112. |
| [171] |
Dragneva Y, Anuradha CD, Valeva A, et al. (2001) Subcytocidal attack by staphylococcal alpha-toxin activates NFkappaB and induces interleukin-8 production. Infect Immun 69: 2630-2635. doi: 10.1128/IAI.69.4.2630-2635.2001
|
| [172] |
Lizak M, Yarovinsky TO (2012) Phospholipid scramblase 1 mediates type I interferon-induced protection against staphylococcal alpha-toxin. Cell Host Microbe 11: 70-80. doi: 10.1016/j.chom.2011.12.004
|
| [173] | la Sala A, Ferrari D, Di Virgilio F, et al. (2003) Alerting and tuning the immune response by extracellular nucle |
| [174] |
Okada SF, Nicholas RA, Kreda SM, et al. (2006) Physiological regulation of ATP release at the apical surface of human airway epithelia. J Biol Chem 281: 22992-23002. doi: 10.1074/jbc.M603019200
|
| [175] | Bond S, Naus CC (2014) The pannexins: Past and present. Front Physiol 5: 1-24. |
| [176] |
Russo MJ, Bayley H, Toner M (1997) Reversible permeabilization of plasma membranes with an engineered switchable pore. Nat Biotechnol 15: 278-282. doi: 10.1038/nbt0397-278
|
| [177] | Cassidy PS, Harshman S (1973) The binding of staphylococcal 125I-alpha-toxin (B) to erythrocytes. J Biol Chem 248: 5545-5546. |
| [178] |
Maurer K, Reyes-Robles T, Alonzo F, 3rd, et al. (2015) Autophagy mediates tolerance to Staphylococcus aureus alpha-toxin. Cell Host Microbe 17: 429-440. doi: 10.1016/j.chom.2015.03.001
|
| [179] |
Kloft N, Neukirch C, Bobkiewicz W, et al. (2010) Pro-autophagic signal induction by bacterial pore-forming toxins. Med Microbiol Immunol 199: 299-309. doi: 10.1007/s00430-010-0163-0
|
| [180] |
Husmann M, Beckmann E, Boller K, et al. (2009) Elimination of a bacterial pore-forming toxin by sequential endocytosis and exocytosis. FEBS Lett 583: 337-344. doi: 10.1016/j.febslet.2008.12.028
|
| [181] |
Kwak YK, Vikstrom E, Magnusson KE, et al. (2012) The Staphylococcus aureus alpha-toxin perturbs the barrier function in Caco-2 epithelial cell monolayers by altering junctional integrity. Infect Immun 80: 1670-1680. doi: 10.1128/IAI.00001-12
|
| [182] |
Boucher RC (2004) New concepts of the pathogenesis of cystic fibrosis lung disease. Eur Respir J 23: 146-158. doi: 10.1183/09031936.03.00057003
|
| [183] |
Kunzelmann K, McMorran B (2004) First encounter: How pathogens compromise epithelial transport. Physiology (Bethesda, Md) 19: 240-244. doi: 10.1152/physiol.00015.2004
|
| [184] |
Tarran R (2004) Regulation of airway surface liquid volume and mucus transport by active ion transport. Proc Am Thoracic Soc 1: 42-46. doi: 10.1513/pats.2306014
|
| [185] |
Escotte S, Al Alam D, Le Naour R, et al. (2006) T cell chemotaxis and chemokine release after Staphylococcus aureus interaction with polarized airway epithelium. Am J Respir Cell Mol Biol 34: 348-354. doi: 10.1165/rcmb.2005-0191OC
|
| [186] |
Lee RJ, Foskett JK (2014) Ca2+ signaling and fluid secretion by secretory cells of the airway epithelium. Cell Calcium 55: 325-336. doi: 10.1016/j.ceca.2014.02.001
|
| [187] | Suttorp N, Hessz T, Seeger W, et al. (1988) Bacterial exotoxins and endothelial permeability for water and albumin in vitro. Am J Physiol Cell Physiol 255: C368-C376. |
| [188] |
Hocke AC, Temmesfeld-Wollbrueck B, Schmeck B, et al. (2006) Perturbation of endothelial junction proteins by Staphylococcus aureus alpha-toxin: Inhibition of endothelial gap formation by adrenomedullin. Histochem Cell Biol 126: 305-316. doi: 10.1007/s00418-006-0174-5
|
| [189] |
Stull JT, Tansey MG, Tang DC, et al. (1993) Phosphorylation of myosin light chain kinase: A cellular mechanism for Ca2+ desensitization. Mol Cell Biochem 127-128: 229-237. doi: 10.1007/BF01076774
|
| [190] | Horiuchi K, Le Gall S, Schulte M, et al. (2007) Substrate selectivity of epidermal growth factor-receptor ligand sheddases and their regulation by phorbol esters and calcium influx. Mol Biol Cell 18: 176-188. |
| [191] |
Le Gall SM, Bobe P, Reiss K, et al. (2009) ADAMs 10 and 17 represent differentially regulated components of a general shedding machinery for membrane proteins such as transforming growth factor alpha, L-selectin, and tumor necrosis factor alpha. Mol Biol Cell 20: 1785-1794. doi: 10.1091/mbc.E08-11-1135
|
| [192] |
Brieher WM, Yap AS (2013) Cadherin junctions and their cytoskeleton(s). Curr Opin Cell Biol 25: 39-46. doi: 10.1016/j.ceb.2012.10.010
|
| [193] |
Zaidel-Bar R, Itzkovitz S, Ma'ayan A, et al. (2007) Functional atlas of the integrin adhesome. Nat Cell Biol 9: 858-867. doi: 10.1038/ncb0807-858
|
| [194] | Dreymueller D, Uhlig S, Ludwig A (2015) ADAM-family metalloproteinases in lung inflammation: Potential therapeutic targets. Am J Physiol 308: L325-L343. |
| [195] |
Sahin U, Weskamp G, Kelly K, et al. (2004) Distinct roles for ADAM10 and ADAM17 in ectodomain shedding of six EGFR ligands. J Cell Biol 164: 769-779. doi: 10.1083/jcb.200307137
|
| [196] |
Park PW, Foster TJ, Nishi E, et al. (2004) Activation of syndecan-1 ectodomain shedding by Staphylococcus aureus alpha-toxin and beta-toxin. J Biol Chem 279: 251-258. doi: 10.1074/jbc.M308537200
|
| [197] |
Hayashida A, Bartlett AH, Foster TJ, et al. (2009) Staphylococcus aureus beta-toxin induces lung injury through syndecan-1. Am J Pathol 174: 509-518. doi: 10.2353/ajpath.2009.080394
|
| [198] |
Tengholm A, Hellman B, Gylfe E (2000) Mobilization of Ca2+ stores in individual pancreatic beta-cells permeabilized or not with digitonin or alpha-toxin. Cell Calcium 27: 43-51. doi: 10.1054/ceca.1999.0087
|
| [199] |
Huang TY, Minamide LS, Bamburg JR, et al. (2008) Chronophin mediates an ATP-sensing mechanism for cofilin dephosphorylation and neuronal cofilin-actin rod formation. Dev Cell 15: 691-703. doi: 10.1016/j.devcel.2008.09.017
|
| [200] |
Eichstaedt S, Gäbler K, Below S, et al. (2008) Phospholipase C-activating plasma membrane receptors and calcium signaling in immortalized human airway epithelial cells. J Recept Signal Transd 28: 591-612. doi: 10.1080/10799890802407120
|
| [201] |
Schwiebert EM, Zsembery A (2003) Extracellular ATP as a signaling molecule for epithelial cells. Biochim Biophys Acta 1615: 7-32. doi: 10.1016/S0005-2736(03)00210-4
|
| [202] |
Tarran R, Button B, Boucher RC (2006) Regulation of normal and cystic fibrosis airway surface liquid volume by phasic shear stress. Annu Rev Physiol 68: 543-561. doi: 10.1146/annurev.physiol.68.072304.112754
|
| [203] |
Evans JH, Sanderson MJ (1999) Intracellular calcium oscillations regulate ciliary beat frequency of airway epithelial cells. Cell Calcium 26: 103-110. doi: 10.1054/ceca.1999.0060
|
| [204] |
Yun YS, Min YG, Rhee CS, et al. (1999) Effects of alpha-toxin of Staphylococcus aureus on the ciliary activity and ultrastructure of human nasal ciliated epithelial cells. Laryngoscope 109: 2021-2024. doi: 10.1097/00005537-199912000-00024
|
| [205] |
Knowles M, Robinson J, Wood R, et al. (1997) Ion composition of airway surface liquid of patients with cystic fibrosis as compared with normal and disease-control subjects. J Clin Invest 100: 2588-2595. doi: 10.1172/JCI119802
|
| [206] |
Olson R, Nariya H, Yokota K, et al. (1999) Crystal structure of staphylococcal LukF delineates conformational changes accompanying formation of a transmembrane channel. Nat Struct Biol 6: 134-140. doi: 10.1038/5821
|
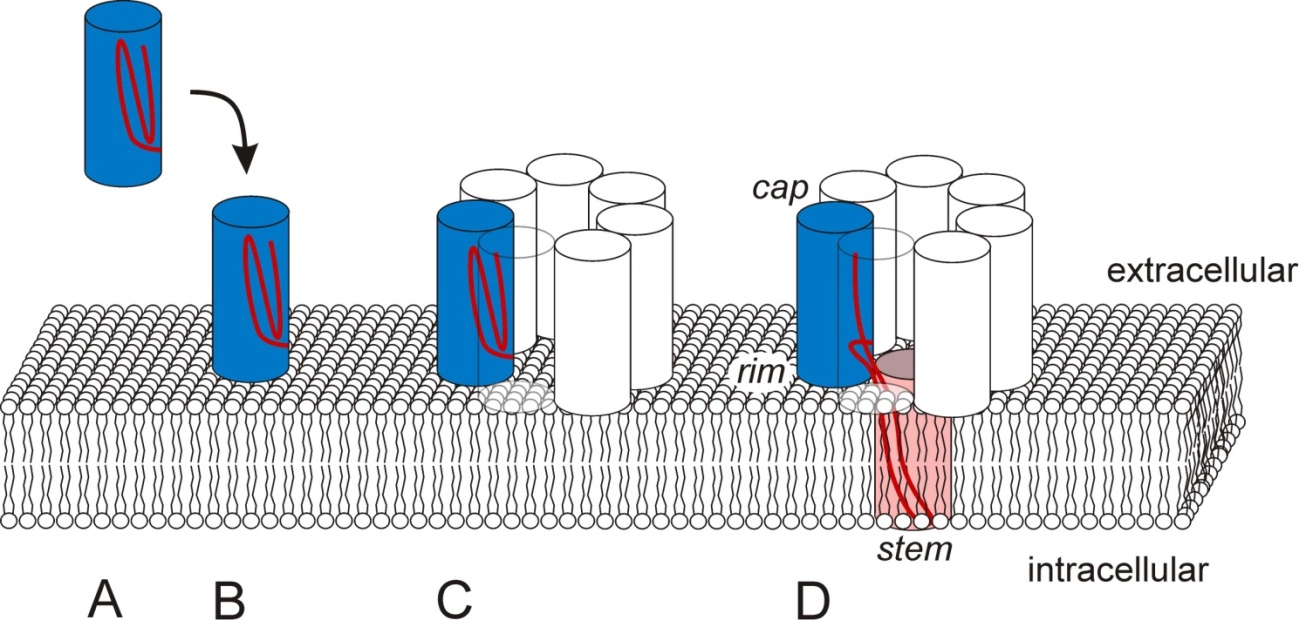
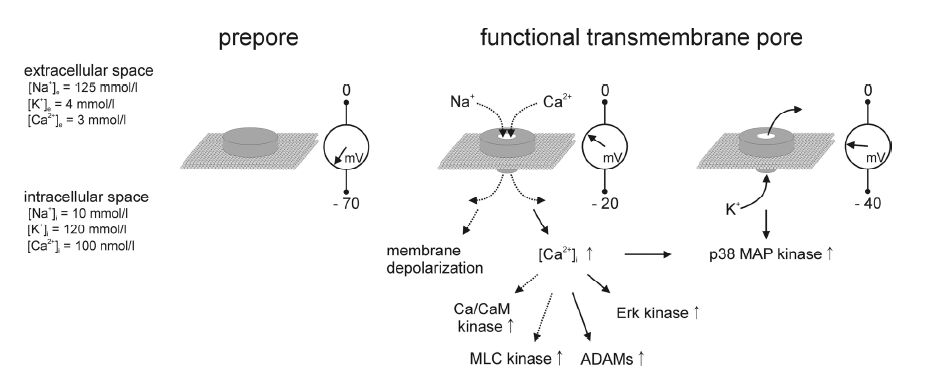
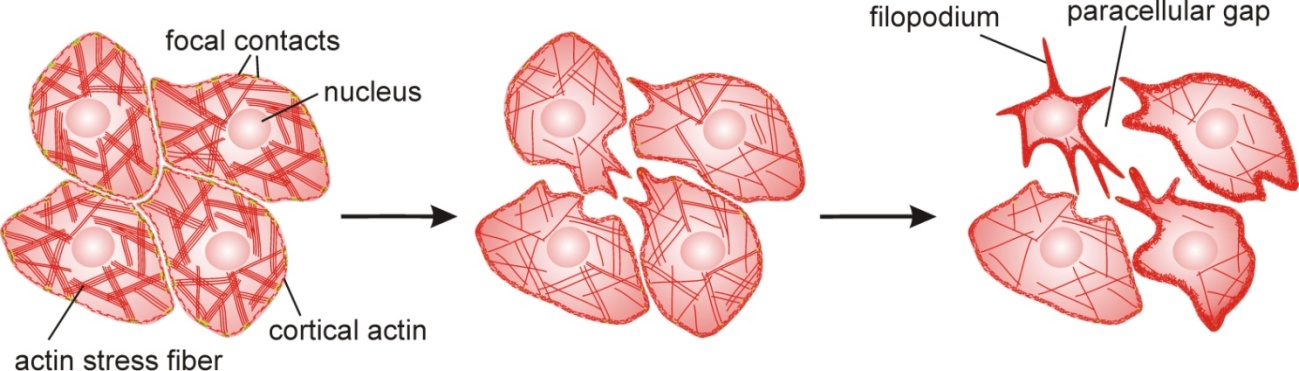
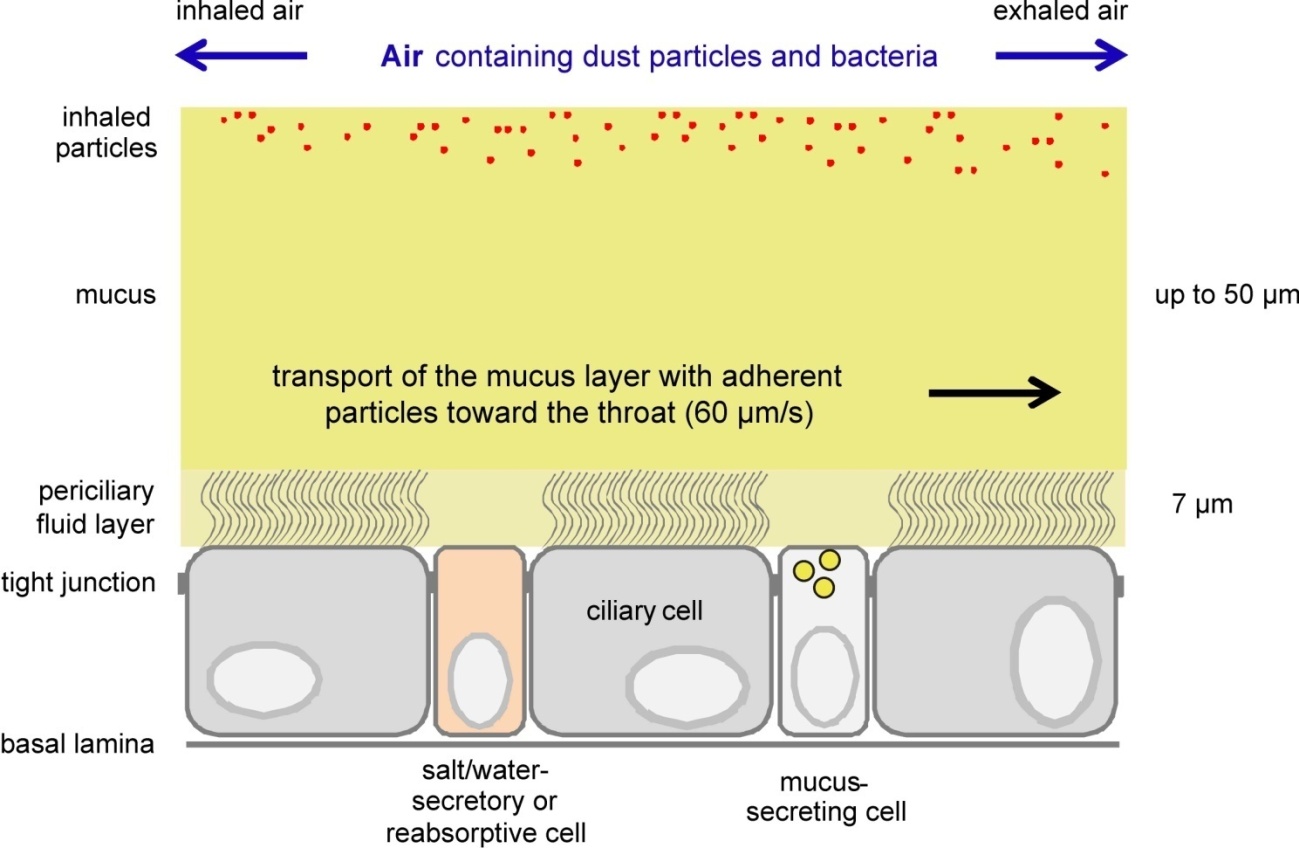
Figures(4)

Jan-Peter Hildebrandt. Pore-forming virulence factors of Staphylococcus aureus destabilize epithelial barriers-effects of alpha-toxin in the early phases of airway infection[J]. AIMS Microbiology, 2015, 1(1): 11-36. doi: 10.3934/microbiol.2015.1.11







 DownLoad:
DownLoad: