Citation: Timothy Jan Bergmann, Giorgia Brambilla Pisoni, Maurizio Molinari. Quality control mechanisms of protein biogenesis: proteostasis dies hard[J]. AIMS Biophysics, 2016, 3(4): 456-478. doi: 10.3934/biophy.2016.4.456

| [1] | Crick FHC (1956) Ideas on protein synthesis. Wellcome Library for the History and Understanding of Medicine. Avaiable from: http://archives.wellcome.ac.uk/. |
| [2] |
Crick FHC (1970) Central Dogma of Molecular Biology. Nature 227: 561–563. doi: 10.1038/227561a0

|
| [3] |
Baltimore D (1970) RNA-dependent DNA polymerase in virions of RNA tumour viruses. Nature 226: 1209–1211. doi: 10.1038/2261209a0
|
| [4] |
Temin HM, Mizutani S (1970) RNA-dependent DNA polymerase in virions of Rous sarcoma virus. Nature 226: 1211–1213. doi: 10.1038/2261211a0
|
| [5] |
Koonin EV (2012) Does the central dogma still stand? Biol Direct 7: 27. doi: 10.1186/1745-6150-7-27
|
| [6] |
Melnikov S, Ben-Shem A, Garreau de Loubresse N, et al. (2012) One core, two shells: bacterial and eukaryotic ribosomes. Nat Struct Mol Biol 19: 560–567. doi: 10.1038/nsmb.2313
|
| [7] |
Khatter H, Myasnikov AG, Natchiar SK, et al. (2015) Structure of the human 80S ribosome. Nature 520: 640–645. doi: 10.1038/nature14427
|
| [8] |
Kolitz SE, Lorsch JR (2010) Eukaryotic initiator tRNA: finely tuned and ready for action. FEBS Lett 584: 396–404. doi: 10.1016/j.febslet.2009.11.047
|
| [9] |
Rodnina MV (2016) The ribosome in action: Tuning of translational efficiency and protein folding. Protein Sci 25: 1390–1406. doi: 10.1002/pro.2950
|
| [10] |
Dabrowski M, Bukowy-Bieryllo Z, Zietkiewicz E (2015) Translational readthrough potential of natural termination codons in eucaryotes--The impact of RNA sequence. RNA Biol 12: 950–958. doi: 10.1080/15476286.2015.1068497
|
| [11] |
Karpinets TV, Greenwood DJ, Sams CE, et al. (2006) RNA:protein ratio of the unicellular organism as a characteristic of phosphorous and nitrogen stoichiometry and of the cellular requirement of ribosomes for protein synthesis. BMC Biol 4: 1–10. doi: 10.1186/1741-7007-4-1
|
| [12] |
Ingolia NT, Lareau LF, Weissman JS (2011) Ribosome profiling of mouse embryonic stem cells reveals the complexity and dynamics of mammalian proteomes. Cell 147: 789–802. doi: 10.1016/j.cell.2011.10.002
|
| [13] |
Dennis PP, Bremer H (1974) Differential Rate of Ribosomal Protein Synthesis in Escherichia coli B/r. J Mol Biol 84: 407–422. doi: 10.1016/0022-2836(74)90449-5
|
| [14] |
Dennis PP, Nomura M (1974) Stringent Control of Ribosomal Protein Gene Expression in Escherichia coli. Proc Nat Acad Sci USA 71: 3819–3823. doi: 10.1073/pnas.71.10.3819
|
| [15] |
Young R, Bremer H (1976) Polypeptide-Chain-Elongation Rate in Escherichia coli B/r as a Function ofGrowth Rate. Biochem J 160: 185–194. doi: 10.1042/bj1600185
|
| [16] | Schaaper RM (1993) Base selection, proofreading, and mismatch repair during DNA replication in Escherichia coli. J Biol Chem 268: 23762–23765. |
| [17] | Drake JW, Charlesworth B, Charlesworth D, et al. (1998) Rates of spontaneous mutation. Genetics 148: 1667–1686. |
| [18] |
Kunkel TA (2004) DNA replication fidelity. J Biol Chem 279: 16895–16898. doi: 10.1074/jbc.R400006200
|
| [19] |
Bebenek K, Kunkel TA (2004) Functions of DNA polymerases. Adv Protein Chem 69: 137–165. doi: 10.1016/S0065-3233(04)69005-X
|
| [20] |
Kunkel TA (2009) Evolving Views of DNA Replication (In)Fidelity. Cold Spring Harb Sym 74: 91–101. doi: 10.1101/sqb.2009.74.027
|
| [21] |
Sainsbury S, Bernecky C, Cramer P (2015) Structural basis of transcription initiation by RNA polymerase II. Nat Rev Mol Cell Biol 16: 129–143. doi: 10.1038/nrm3952
|
| [22] |
Sharma N (2016) Regulation of RNA polymerase II-mediated transcriptional elongation: Implications in human disease. IUBMB Life 68: 709–716. doi: 10.1002/iub.1538
|
| [23] | Loya TJ, Reines D (2016) Recent advances in understanding transcription termination by RNA polymerase II. F1000 Res 5: 1478. |
| [24] |
Schwanhausser B, Busse D, Li N, et al. (2011) Global quantification of mammalian gene expression control. Nature 473: 337–342. doi: 10.1038/nature10098
|
| [25] |
Imashimizu M, Oshima T, Lubkowska L, et al. (2013) Direct assessment of transcription fidelity by high-resolution RNA sequencing. Nucleic Acids Res 41: 9090–9104. doi: 10.1093/nar/gkt698
|
| [26] |
Ninio J (1991) Connections between translation, transcription and replication error-rates. Biochimie 73: 1517–1523. doi: 10.1016/0300-9084(91)90186-5
|
| [27] |
Gouta JF, Thomasb WK, Smithc Z, et al. (2013) Large-scale detection of in vivo transcription errors. PNAS 110: 18584–18589. doi: 10.1073/pnas.1309843110
|
| [28] |
Cochella L, Green R (2005) Fidelity in protein synthesis. Curr Biol 15: 536–540. doi: 10.1016/j.cub.2005.02.019
|
| [29] | Kramer EB, Farabaugh PJ (2007) The frequency of translational misreading errors in E. coli is largely determined by tRNA competition. RNA 13: 87–96. |
| [30] |
Zaher HS, Green R (2009) Fidelity at the molecular level: lessons from protein synthesis. Cell 136: 746–762. doi: 10.1016/j.cell.2009.01.036
|
| [31] | Gingold H, Pilpel Y (2011) Determinants of translation efficiency and accuracy. Mol Syst Biol 7: 141–150. |
| [32] |
Ribas de Pouplana L, Santos MA, Zhu JH, et al. (2014) Protein mistranslation: friend or foe? Trends Biochem Sci 39: 355–362. doi: 10.1016/j.tibs.2014.06.002
|
| [33] |
Anfinsen CB (1973) Principles that govern the folding of protein chains. Science 181: 223–230. doi: 10.1126/science.181.4096.223
|
| [34] |
Bulik S, Peters B, Holzhutter HG (2005) Quantifying the Contribution of Defective Ribosomal Products to Antigen Production: A Model-Based Computational Analysis. J Immunol 175: 7957–7964. doi: 10.4049/jimmunol.175.12.7957
|
| [35] |
Vabulas RM, Hartl FU (2005) Protein synthesis upon acute nutrient restriction relies on proteasome function. Science 310: 1960–1963. doi: 10.1126/science.1121925
|
| [36] |
Schubert U, Antón LC, Gibbs J, et al. (2000) Rapid degradation of a large fraction of newly synthesized proteins by proteasomes. Nature 404: 770–774. doi: 10.1038/35008096
|
| [37] |
Vabulas RM, Hartl UF (2005) Protein Synthesis upon Acute Nutrient Restriction Relies on Proteasome Function. Science 310: 1960–1963. doi: 10.1126/science.1121925
|
| [38] | Hung MC, Link W (2011) Protein localization in disease and therapy. J Cell Sci 124: 3381–3392. |
| [39] |
Chacinska A, Koehler CM, Milenkovic D, et al. (2009) Importing mitochondrial proteins: machineries and mechanisms. Cell 138: 628–644. doi: 10.1016/j.cell.2009.08.005
|
| [40] |
Rapoport TA (2007) Protein translocation across the eukaryotic endoplasmic reticulum and bacterial plasma membranes. Nature 450: 663–669. doi: 10.1038/nature06384
|
| [41] |
Geva Y, Schuldiner M (2014) The back and forth of cargo exit from the endoplasmic reticulum. Curr Biol 24: 130–136. doi: 10.1016/j.cub.2013.12.008
|
| [42] |
Barlowe C, Helenius A (2016) Cargo Capture and Bulk Flow in the Early Secretory Pathway. Annu Rev Cell Dev Biol 32: 197–222. doi: 10.1146/annurev-cellbio-111315-125016
|
| [43] |
Herrmann JM, Neupert W (2000) Protein transport into mitochondria. Curr Opin Microbiol 3: 210–214. doi: 10.1016/S1369-5274(00)00077-1
|
| [44] |
Zimmermann R, Eyrisch S, Ahmad M, et al. (2011) Protein translocation across the ER membrane. Biochim Biophys Acta 1808: 912–924. doi: 10.1016/j.bbamem.2010.06.015
|
| [45] |
Freitas N, Cunha C (2009) Mechanisms and signals for the nuclear import of proteins. Curr Genomics 10: 550–557. doi: 10.2174/138920209789503941
|
| [46] |
Nichols WC, Seligsohn U, Zivelin A, et al. (1998) Mutations in the ER-Golgi intermediate compartment protein ERGIC-53 cause combined deficiency of coagulation factors V and VIII. Cell 93: 61–70. doi: 10.1016/S0092-8674(00)81146-0
|
| [47] |
Spreafico M, Peyvandi F (2009) Combined Factor V and Factor VIII Deficiency. Semin Thromb Hemost 35: 390–399. doi: 10.1055/s-0029-1225761
|
| [48] |
Rock KL, Gramm C, Rothstein L, et al. (1994) Inhibitors of the proteasome block the degradation of most cell proteins and the generation of peptides presented on MHC class I molecules. Cell 78: 761–771. doi: 10.1016/S0092-8674(94)90462-6
|
| [49] |
Goldberg AL (2003) Protein degradation and protection against misfolded or damaged proteins. Nature 426: 895–899. doi: 10.1038/nature02263
|
| [50] |
Finley D (2009) Recognition and processing of ubiquitin-protein conjugates by the proteasome. Annu Rev Biochem 78: 477–513. doi: 10.1146/annurev.biochem.78.081507.101607
|
| [51] |
Rock KL, Farfan-Arribas DJ, Colbert JD, et al. (2014) Re-examining class-I presentation and the DRiP hypothesis. Trends Immunol 35: 144–152. doi: 10.1016/j.it.2014.01.002
|
| [52] | Cohen-Kaplan V, Livneh I, Avni N, et al. (2016) The ubiquitin-proteasome system and autophagy: Coordinated and independent activities. Int J Biochem Cell Biol: In Press. |
| [53] |
Ravikumar B, Sarkar S, Davies JE, et al. (2010) Regulation of mammalian autophagy in physiology and pathophysiology. Physiol Rev 90: 1383–1435. doi: 10.1152/physrev.00030.2009
|
| [54] |
Mariappan M, Li X, Stefanovic S, et al. (2010) A ribosome-associating factor chaperones tail-anchored membrane proteins. Nature 466: 1120–1124. doi: 10.1038/nature09296
|
| [55] |
Brandman O, Stewart-Ornstein J, Wong D, et al. (2012) A ribosome-bound quality control complex triggers degradation of nascent peptides and signals translation stress. Cell 151: 1042–1054. doi: 10.1016/j.cell.2012.10.044
|
| [56] |
Defenouillère Q, Yao Y, Mouaikel J, et al. (2013) Cdc48 associated complex bound to 60s particles is required for the clearance of aberrant translation products. PNAS 110: 5046–5051. doi: 10.1073/pnas.1221724110
|
| [57] |
Shao S, von der Malsburg K, Hegde RS (2013) Listerin-dependent nascent protein ubiquitination relies on ribosome subunit dissociation. Mol Cell 50: 637–648. doi: 10.1016/j.molcel.2013.04.015
|
| [58] | Verma R, Oania RS, Kolawa1 NJ, et al. (2013) Cdc48/p97 promotes degradation of aberrant nascent polypeptides bound to the ribosome. eLife 2: e00308. |
| [59] | Shen PS, Park J, Qin Y, et al. (2016) Rqc2p and 60S ribosomal subunits mediate mRNA-independent elongation of nascent chains. Science 347: 75–78. |
| [60] |
Ghaemmaghami S, Huh WK, Bower K, et al. (2003) Global analysis of protein expression in yeast. Nature 425: 737–741. doi: 10.1038/nature02046
|
| [61] |
Schwarz F, Aebi M (2011) Mechanisms and principles of N-linked protein glycosylation. Curr Opin Struct Biol 21: 576–582. doi: 10.1016/j.sbi.2011.08.005
|
| [62] |
Tannous A, Pisoni GB, Hebert DN, et al. (2015) N-linked sugar-regulated protein folding and quality control in the ER. Semin Cell Dev Biol 41: 79–89. doi: 10.1016/j.semcdb.2014.12.001
|
| [63] |
Ellgaard L, Molinari M, Helenius A (1999) Setting the standards: quality control in the secretory pathway. Science 286: 1882–1888. doi: 10.1126/science.286.5446.1882
|
| [64] |
Aebi M, Bernasconi R, Clerc S, et al. (2010) N-glycan structures: recognition and processing in the ER. Trends Biochem Sci 35: 74–82. doi: 10.1016/j.tibs.2009.10.001
|
| [65] |
Schallus T, Feher K, Sternberg U, et al. (2010) Analysis of the specific interactions between the lectin domain of malectin and diglucosides. Glycobiology 20: 1010–1020. doi: 10.1093/glycob/cwq059
|
| [66] |
Galli C, Bernasconi R, Solda T, et al. (2011) Malectin participates in a backup glycoprotein quality control pathway in the mammalian ER. PLoS One 6: e16304. doi: 10.1371/journal.pone.0016304
|
| [67] |
Pisoni GB, Ruddock LW, Bulleid N, et al. (2015) Division of labor among oxidoreductases: TMX1 preferentially acts on transmembrane polypeptides. Mol Biol Cell 26: 3390–3400. doi: 10.1091/mbc.E15-05-0321
|
| [68] |
Lamriben L, Graham JB, Adams BM, et al. (2016) N-Glycan-based ER Molecular Chaperone and Protein Quality Control System: The Calnexin Binding Cycle. Traffic 17: 308–326. doi: 10.1111/tra.12358
|
| [69] |
Cabral CM, Choudhury P, Liu Y, et al. (2000) Processing by endoplasmic reticulum mannosidases partitions a secretion-impaired glycoprotein into distinct disposal pathways. J Biol Chem 275: 25015–25022. doi: 10.1074/jbc.M910172199
|
| [70] |
Olivari S, Cali T, Salo KE, et al. (2006) EDEM1 regulates ER-associated degradation by accelerating de-mannosylation of folding-defective polypeptides and by inhibiting their covalent aggregation. Biochem Biophys Res Commun 349: 1278–1284. doi: 10.1016/j.bbrc.2006.08.186
|
| [71] |
Ninagawa S, Okada T, Sumitomo Y, et al. (2014) EDEM2 initiates mammalian glycoprotein ERAD by catalyzing the first mannose trimming step. J Cell Biol 206: 347–356. doi: 10.1083/jcb.201404075
|
| [72] |
Hirao K, Natsuka Y, Tamura T, et al. (2006) EDEM3, a soluble EDEM homolog, enhances glycoprotein endoplasmic reticulum-associated degradation and mannose trimming. J Biol Chem 281: 9650–9658. doi: 10.1074/jbc.M512191200
|
| [73] |
Olivari S, Molinari M (2007) Glycoprotein folding and the role of EDEM1, EDEM2 and EDEM3 in degradation of folding-defective glycoproteins. FEBS Lett 581: 3658–3664. doi: 10.1016/j.febslet.2007.04.070
|
| [74] |
Christianson JC, Shaler TA, Tyler RE, et al. (2008) OS-9 and GRP94 deliver mutant alpha1-antitrypsin to the Hrd1-SEL1L ubiquitin ligase complex for ERAD. Nat Cell Biol 10: 272–282. doi: 10.1038/ncb1689
|
| [75] |
Bernasconi R, Galli C, Calanca V, et al. (2010) Stringent requirement for HRD1, SEL1L, and OS-9/XTP3-B for disposal of ERAD-LS substrates. J Cell Biol 188: 223–235. doi: 10.1083/jcb.200910042
|
| [76] |
Vembar SS, Brodsky JL (2008) One step at a time: endoplasmic reticulum-associated degradation. Nat Rev Mol Cell Biol 9: 944–957. doi: 10.1038/nrm2546
|
| [77] |
Merulla J, Solda T, Molinari M (2015) A novel UGGT1 and p97-dependent checkpoint for native ectodomains with ionizable intramembrane residue. Mol Biol Cell 26: 1532–1542. doi: 10.1091/mbc.E14-12-1615
|
| [78] |
Merulla J, Fasana E, Solda T, et al. (2013) Specificity and regulation of the endoplasmic reticulum-associated degradation machinery. Traffic 14: 767–777. doi: 10.1111/tra.12068
|
| [79] |
Bernasconi R, Molinari M (2011) ERAD and ERAD tuning: disposal of cargo and of ERAD regulators from the mammalian ER. Curr Opin Cell Biol 23: 176–183. doi: 10.1016/j.ceb.2010.10.002
|
| [80] |
Koenig PA, Nicholls PK, Schmidt FI, et al. (2014) The E2 ubiquitin-conjugating enzyme UBE2J1 is required for spermiogenesis in mice. J Biol Chem 289: 34490–34502. doi: 10.1074/jbc.M114.604132
|
| [81] |
Hagiwara M, Ling J, Koenig PA, et al. (2016) Posttranscriptional Regulation of Glycoprotein Quality Control in the Endoplasmic Reticulum Is Controlled by the E2 Ub-Conjugating Enzyme UBC6e. Mol Cell 63: 753–767. doi: 10.1016/j.molcel.2016.07.014
|
| [82] |
Glick D, Barth S, Macleod KF (2010) Autophagy: cellular and molecular mechanisms. J Pathol 221: 3–12. doi: 10.1002/path.2697
|
| [83] |
Mizushima N, Ohsumi Y, Yoshimori T (2002) Autophagosome Formation in Mammalian Cells. Cell Struct Funct 27: 421–429. doi: 10.1247/csf.27.421
|
| [84] |
Ohsumi Y (2014) Historical landmarks of autophagy research. Cell Res 24: 9–23. doi: 10.1038/cr.2013.169
|
| [85] | Ariosa AR, Klionsky DJ (2016) Autophagy core machinery: overcoming spatial barriers in neurons. J Mol Med (Berl): In Press. |
| [86] |
Ryter SW, Cloonan SM, Choi AM (2013) Autophagy: a critical regulator of cellular metabolism and homeostasis. Mol Cells 36: 7–16. doi: 10.1007/s10059-013-0140-8
|
| [87] |
Choi AM, Ryter SW, Levine B (2013) Autophagy in human health and disease. N Engl J Med 368: 1845–1846. doi: 10.1056/NEJMc1303158
|
| [88] | Lin F, Qin ZH (2013) Degradation of misfolded proteins by autophagy: is it a strategy for Huntington's disease treatment? J Huntingtons Dis 2: 149–157. |
| [89] |
Webb JL, Ravikumar B, Atkins J, et al. (2003) Alpha-Synuclein is degraded by both autophagy and the proteasome. J Biol Chem 278: 25009–25013. doi: 10.1074/jbc.M300227200
|
| [90] | Pickford F, Masliah E, Britschgi M, et al. (2008) The autophagy-related protein beclin 1 shows reduced expression in early Alzheimer disease and regulates amyloid beta accumulation in mice. J Clin Invest 118: 2190–2199. |
| [91] |
Lee MJ, Lee JH, Rubinsztein DC (2013) Tau degradation: the ubiquitin-proteasome system versus the autophagy-lysosome system. Prog Neurobiol 105: 49–59. doi: 10.1016/j.pneurobio.2013.03.001
|
| [92] |
Perlmutter DH (2011) Alpha-1-antitrypsin deficiency: importance of proteasomal and autophagic degradative pathways in disposal of liver disease-associated protein aggregates. Annu Rev Med 62: 333–345. doi: 10.1146/annurev-med-042409-151920
|
| [93] |
Fu L, Sztul E (2009) ER-associated complexes (ERACs) containing aggregated cystic fibrosis transmembrane conductance regulator (CFTR) are degraded by autophagy. Eur J Cell Biol 88: 215–226. doi: 10.1016/j.ejcb.2008.11.003
|
| [94] | Farre JC, Subramani S (2016) Mechanistic insights into selective autophagy pathways: lessons from yeast. Nat Rev Mol Cell Biol 17: 537–552. |
| [95] |
Khaminets A, Heinrich T, Mari M, et al. (2015) Regulation of endoplasmic reticulum turnover by selective autophagy. Nature 522: 354–358. doi: 10.1038/nature14498
|
| [96] | Fumagalli FNJ, Bergmann TJ, Cebollero E, et al. (2016) Translocon component Sec62 acts in endoplasmic reticulum turnover during stress recovery. Nat Cell Biol: In press. |
| [97] |
Inoue T, Tsai B (2013) How viruses use the endoplasmic reticulum for entry, replication, and assembly. Cold Spring Harb Perspect Biol 5: a013250. doi: 10.1101/cshperspect.a013250
|
| [98] |
van den Boomen DJ, Lehner PJ (2015) Identifying the ERAD ubiquitin E3 ligases for viral and cellular targeting of MHC class I. Mol Immunol 68: 106–111. doi: 10.1016/j.molimm.2015.07.005
|
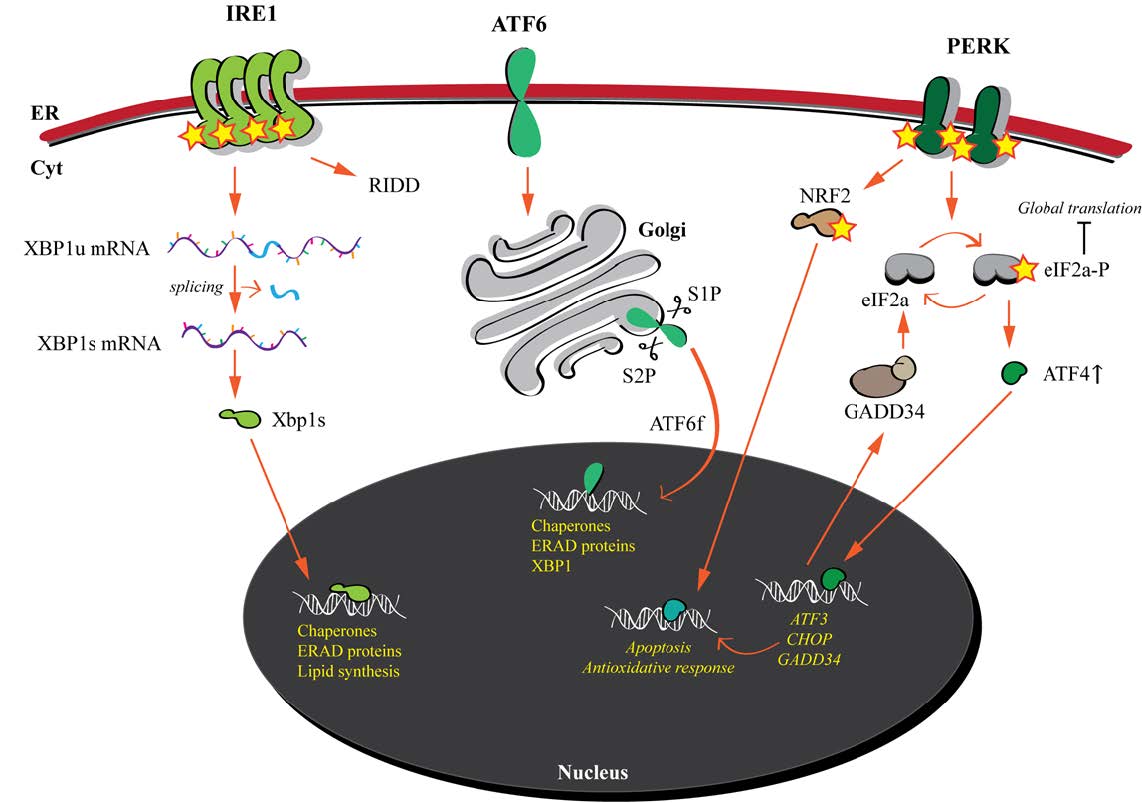
| [99] | Gardner BM, Pincus D, Gotthardt K, et al. (2013) Endoplasmic reticulum stress sensing in the unfolded protein response. Cold Spring Harb Perspect Biol 5: a013169. |
| [100] |
Mori K (2009) Signalling pathways in the unfolded protein response: development from yeast to mammals. J Biochem 146: 743–750. doi: 10.1093/jb/mvp166
|
| [101] |
Zhang L, Zhang C, Wang A (2016) Divergence and Conservation of the Major UPR Branch IRE1-bZIP Signaling Pathway across Eukaryotes. Sci Rep 6: 27362. doi: 10.1038/srep27362
|
| [102] |
Lee AH, Iwakoshi NN, Glimcher LH (2003) XBP-1 regulates a subset of endoplasmic reticulum resident chaperone genes in the unfolded protein response. Mol Cell Biol 23: 7448–7459. doi: 10.1128/MCB.23.21.7448-7459.2003
|
| [103] |
Hollien J, Weissman JS (2006) Decay of endoplasmic reticulum-localized mRNAs during the unfolded protein response. Science 313: 104–107. doi: 10.1126/science.1129631
|
| [104] |
Haze K, Yoshida H, Yanagi H, et al. (1999) Mammalian transcription factor ATF6 is synthesized as a transmembrane protein and activated by proteolysis in response to endoplasmic reticulum stress. Mol Biol Cell 10: 3787–3799. doi: 10.1091/mbc.10.11.3787
|
| [105] |
Shoulders MD, Ryno LM, Genereux JC, et al. (2013) Stress-independent activation of XBP1s and/or ATF6 reveals three functionally diverse ER proteostasis environments. Cell Rep 3: 1279–1292. doi: 10.1016/j.celrep.2013.03.024
|
| [106] |
Bertolotti A, Zhang Y, Hendershot LM, et al. (2000) Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat Cell Biol 2: 326–332. doi: 10.1038/35014014
|
| [107] |
Harding HP, Zhang Y, Ron D (1999) Protein translation and folding are coupled by an endoplasmic-reticulum-resident kinase. Nature 397: 271–274. doi: 10.1038/16729
|
| [108] |
Vattem KM, Wek RC (2004) Reinitiation involving upstream ORFs regulates ATF4 mRNA translation in mammalian cells. Proc Natl Acad Sci U S A 101: 11269–11274. doi: 10.1073/pnas.0400541101
|
| [109] |
Lu PD, Harding HP, Ron D (2004) Translation reinitiation at alternative open reading frames regulates gene expression in an integrated stress response. J Cell Biol 167: 27–33. doi: 10.1083/jcb.200408003
|
| [110] |
Jiang HY, Wek SA, McGrath BC, et al. (2004) Activating transcription factor 3 is integral to the eukaryotic initiation factor 2 kinase stress response. Mol Cell Biol 24: 1365–1377. doi: 10.1128/MCB.24.3.1365-1377.2004
|
| [111] |
Schroder M, Kaufman RJ (2005) ER stress and the unfolded protein response. Mutat Res 569: 29–63. doi: 10.1016/j.mrfmmm.2004.06.056
|
| [112] |
Redler RL, Das J, Diaz JR, et al. (2016) Protein Destabilization as a Common Factor in Diverse Inherited Disorders. J Mol Evol 82: 11–16. doi: 10.1007/s00239-015-9717-5
|
| [113] |
Sitia R, Braakman I (2003) Quality control in the endoplasmic reticulum protein factory. Nature 426: 891–894. doi: 10.1038/nature02262
|
| [114] |
Bernier V, Lagace M, Bichet DG, et al. (2004) Pharmacological chaperones: potential treatment for conformational diseases. Trends Endocrinol Metab 15: 222–228. doi: 10.1016/j.tem.2004.05.003
|
| [115] |
Molinari M (2007) N-glycan structure dictates extension of protein folding or onset of disposal. Nat Chem Biol 3: 313–320. doi: 10.1038/nchembio880
|
| [116] |
Kopito RR, Ron D (2000) Conformational disease. Nat Cell Biol 2: 207–209. doi: 10.1038/35041139
|
| [117] |
Gidalevitz T, Ben-Zvi A, Ho KH, et al. (2006) Progressive disruption of cellular protein folding in models of polyglutamine diseases. Science 311: 1471–1474. doi: 10.1126/science.1124514
|
| [118] |
Powers ET, Morimoto RI, Dillin A, et al. (2009) Biological and chemical approaches to diseases of proteostasis deficiency. Annu Rev Biochem 78: 959–991. doi: 10.1146/annurev.biochem.052308.114844
|
| [119] |
Morello JP, Petaja-Repo UE, Bichet DG, et al. (2000) Pharmacological chaperones: a new twist on receptor folding. Trends Pharmacol Sci 21: 466–469. doi: 10.1016/S0165-6147(00)01575-3
|
| [120] |
Cohen FE, Kelly JW (2003) Therapeutic approaches to protein-misfolding diseases. Nature 426: 905–909. doi: 10.1038/nature02265
|
| [121] |
Convertino M, Das J, Dokholyan NV (2016) Pharmacological Chaperones: Design and Development of New Therapeutic Strategies for the Treatment of Conformational Diseases. ACS Chem Biol 11: 1471–1489. doi: 10.1021/acschembio.6b00195
|
| [122] |
Ma Y, Hendershot LM (2004) The role of the unfolded protein response in tumour development: friend or foe? Nat Rev Cancer 4: 966–977. doi: 10.1038/nrc1505
|
| [123] |
Fernandez PM, Tabbara SO, Jacobs LK, et al. (2000) Overexpression of the glucose-regulated stress gene GRP78 in malignant but not benign human breast lesions. Breast Cancer Res Treat 59: 15–26. doi: 10.1023/A:1006332011207
|
| [124] | Shuda M, Kondoh N, Imazeki N, et al. (2003) Activation of the ATF6, XBP1 and grp78 genes in human hepatocellular carcinoma: a possible involvement of the ER stress pathway in hepatocarcinogenesis. J Hepatol 38: 605–614. |
| [125] | Song MS, Park YK, Lee JH, et al. (2001) Induction of glucose-regulated protein 78 by chronic hypoxia in human gastric tumor cells through a protein kinase C-epsilon/ERK/AP-1 signaling cascade. Cancer Res 61: 8322–8330. |
| [126] |
Gazit G, Lu J, Lee AS (1999) De-regulation of GRP stress protein expression in human breast cancer cell lines. Breast Cancer Res Treat 54: 135–146. doi: 10.1023/A:1006102411439
|
| [127] | Plate L, Paxman RJ, Wiseman RL, et al. (2016) Modulating protein quality control. Elife 5: e18431. |
| [128] |
Wang X, Venable J, LaPointe P, et al. (2006) Hsp90 cochaperone Aha1 downregulation rescues misfolding of CFTR in cystic fibrosis. Cell 127: 803–815. doi: 10.1016/j.cell.2006.09.043
|
| [129] |
Mu TW, Ong DS, Wang YJ, et al. (2008) Chemical and biological approaches synergize to ameliorate protein-folding diseases. Cell 134: 769–781. doi: 10.1016/j.cell.2008.06.037
|
| [130] |
Chiang WC, Hiramatsu N, Messah C, et al. (2012) Selective activation of ATF6 and PERK endoplasmic reticulum stress signaling pathways prevent mutant rhodopsin accumulation. Invest Ophthalmol Vis Sci 53: 7159–7166. doi: 10.1167/iovs.12-10222
|
| [131] |
Luheshi LM, Dobson CM (2009) Bridging the gap: from protein misfolding to protein misfolding diseases. FEBS Lett 583: 2581–2586. doi: 10.1016/j.febslet.2009.06.030
|
| [132] |
Braakman I, Bulleid NJ (2011) Protein folding and modification in the mammalian endoplasmic reticulum. Annu Rev Biochem 80: 71–99. doi: 10.1146/annurev-biochem-062209-093836
|
| [133] |
Brodsky JL, Skach WR (2011) Protein folding and quality control in the endoplasmic reticulum: Recent lessons from yeast and mammalian cell systems. Curr Opin Cell Biol 23: 464–475. doi: 10.1016/j.ceb.2011.05.004
|
| [134] |
Papa FR, Zhang C, Shokat K, et al. (2003) Bypassing a kinase activity with an ATP-competitive drug. Science 302: 1533–1537. doi: 10.1126/science.1090031
|
| [135] |
Wiseman RL, Zhang Y, Lee KP, et al. (2010) Flavonol activation defines an unanticipated ligand-binding site in the kinase-RNase domain of IRE1. Mol Cell 38: 291–304. doi: 10.1016/j.molcel.2010.04.001
|
| [136] |
Wang L, Perera BG, Hari SB, et al. (2012) Divergent allosteric control of the IRE1alpha endoribonuclease using kinase inhibitors. Nat Chem Biol 8: 982–989. doi: 10.1038/nchembio.1094
|
| [137] | Sidrauski C, Tsai JC, Kampmann M, et al. (2015) Pharmacological dimerization and activation of the exchange factor eIF2B antagonizes the integrated stress response. Elife 4: e07314. |
| [138] |
Robblee MM, Kim CC, Porter Abate J, et al. (2016) Saturated Fatty Acids Engage an IRE1alpha-Dependent Pathway to Activate the NLRP3 Inflammasome in Myeloid Cells. Cell Rep 14: 2611–2623. doi: 10.1016/j.celrep.2016.02.053
|
| [139] | Gallagher CM, Walter P (2016) Ceapins inhibit ATF6alpha signaling by selectively preventing transport of ATF6alpha to the Golgi apparatus during ER stress. Elife 5: e11880. |
| [140] | Gallagher CM, Garri C, Cain EL, et al. (2016) Ceapins are a new class of unfolded protein response inhibitors, selectively targeting the ATF6alpha branch. Elife 5: e11880. |
| [141] | Plate L, Cooley CB, Chen JJ, et al. (2016) Small molecule proteostasis regulators that reprogram the ER to reduce extracellular protein aggregation. Elife 5: e15550. |
| [142] |
Hsu PD, Lander ES, Zhang F (2014) Development and applications of CRISPR-Cas9 for genome engineering. Cell 157: 1262–1278. doi: 10.1016/j.cell.2014.05.010
|
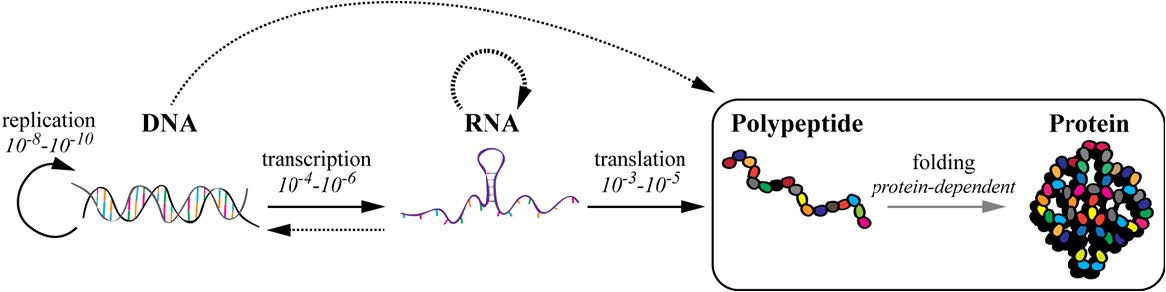
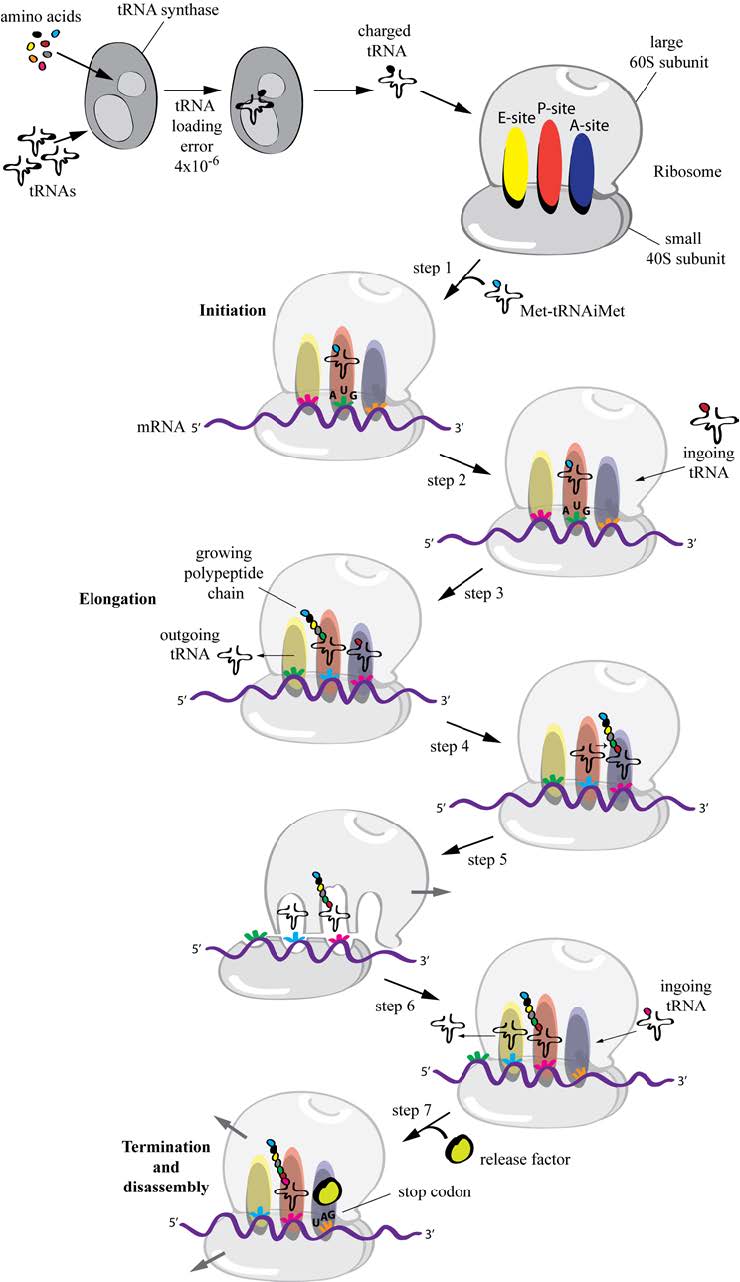
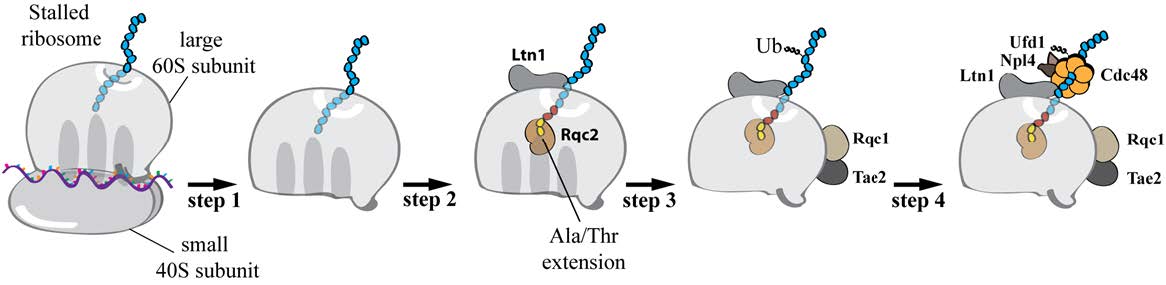
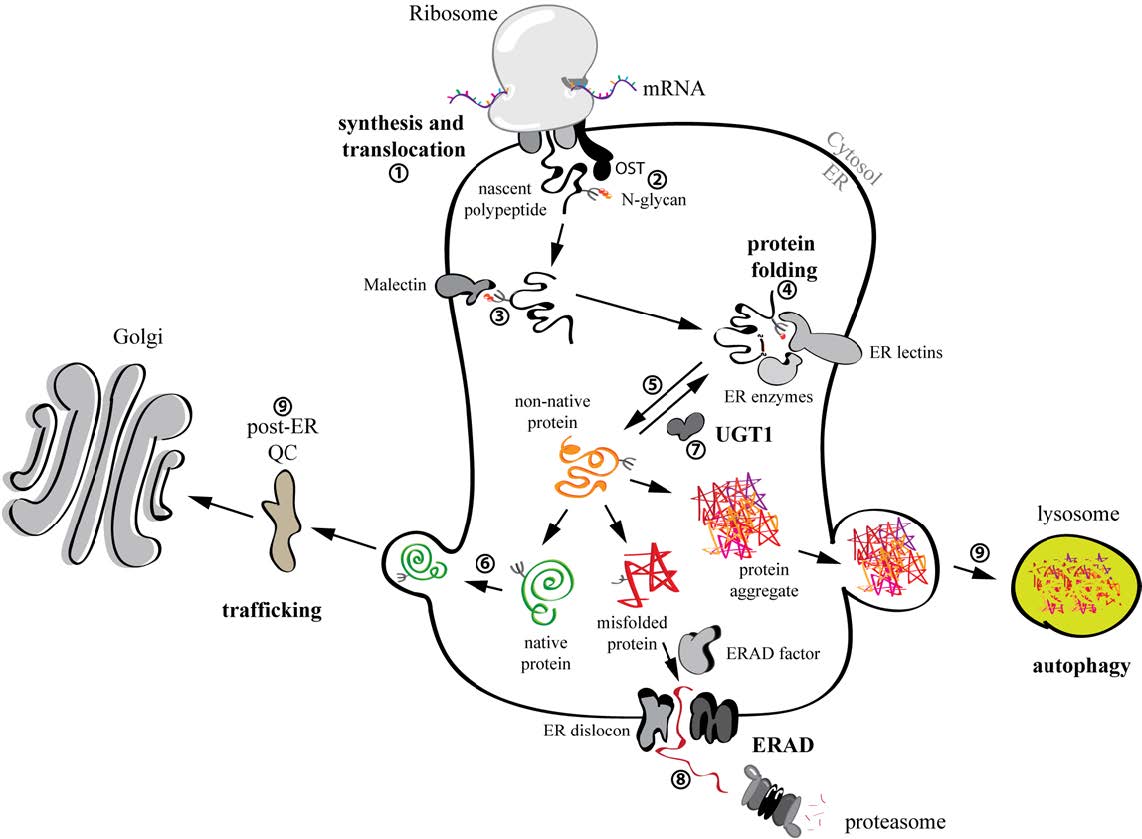
Figures(5)

Timothy Jan Bergmann, Giorgia Brambilla Pisoni, Maurizio Molinari. Quality control mechanisms of protein biogenesis: proteostasis dies hard[J]. AIMS Biophysics, 2016, 3(4): 456-478. doi: 10.3934/biophy.2016.4.456







 DownLoad:
DownLoad: