Citation: Wei Zhang, Sheng Cao, Jessica L. Martin, Joachim D. Mueller, Louis M. Mansky. Morphology and ultrastructure of retrovirus particles[J]. AIMS Biophysics, 2015, 2(3): 343-369. doi: 10.3934/biophy.2015.3.343
| [1] | Cunyi Yang, Li Chen, Bin Mo . The spillover effect of international monetary policy on China's financial market. Quantitative Finance and Economics, 2023, 7(4): 508-537. doi: 10.3934/QFE.2023026 |
| [2] | Raéf Bahrini, Assaf Filfilan . Impact of the novel coronavirus on stock market returns: evidence from GCC countries. Quantitative Finance and Economics, 2020, 4(4): 640-652. doi: 10.3934/QFE.2020029 |
| [3] | Juan David Vega Baquero, Miguel Santolino . Capital flows in integrated capital markets: MILA case. Quantitative Finance and Economics, 2022, 6(4): 622-639. doi: 10.3934/QFE.2022027 |
| [4] | Nan Zhang, Li Zhu . Structural changes in China's flow of funds (1992–2022): A Who-to-Whom model. Quantitative Finance and Economics, 2025, 9(1): 167-201. doi: 10.3934/QFE.2025006 |
| [5] | Arifenur Güngör, Hüseyin Taştan . On macroeconomic determinants of co-movements among international stock markets: evidence from DCC-MIDAS approach. Quantitative Finance and Economics, 2021, 5(1): 19-39. doi: 10.3934/QFE.2021002 |
| [6] | Kim Hiang Liow, Jeongseop Song, Xiaoxia Zhou . Volatility connectedness and market dependence across major financial markets in China economy. Quantitative Finance and Economics, 2021, 5(3): 397-420. doi: 10.3934/QFE.2021018 |
| [7] | Dumisani Pamba, Sophia Mukorera, Peter Moores-Pitt . The asymmetric effects of cross-border equity flow volatility on equity market returns in SANEK countries. Quantitative Finance and Economics, 2025, 9(1): 40-75. doi: 10.3934/QFE.2025002 |
| [8] | Tinghui Li, Jiehua Ma . Does digital finance benefit the income of rural residents? A case study on China. Quantitative Finance and Economics, 2021, 5(4): 664-688. doi: 10.3934/QFE.2021030 |
| [9] | Mustafa Tevfik Kartal, Özer Depren, Serpil Kılıç Depren . The determinants of main stock exchange index changes in emerging countries: evidence from Turkey in COVID-19 pandemic age. Quantitative Finance and Economics, 2020, 4(4): 526-541. doi: 10.3934/QFE.2020025 |
| [10] | Tchai Tavor . Exploring the heterogeneity of stock market responses to Uber announcements: A comparative analysis of developed and emerging economies in Asia Pacific. Quantitative Finance and Economics, 2024, 8(2): 315-346. doi: 10.3934/QFE.2024012 |
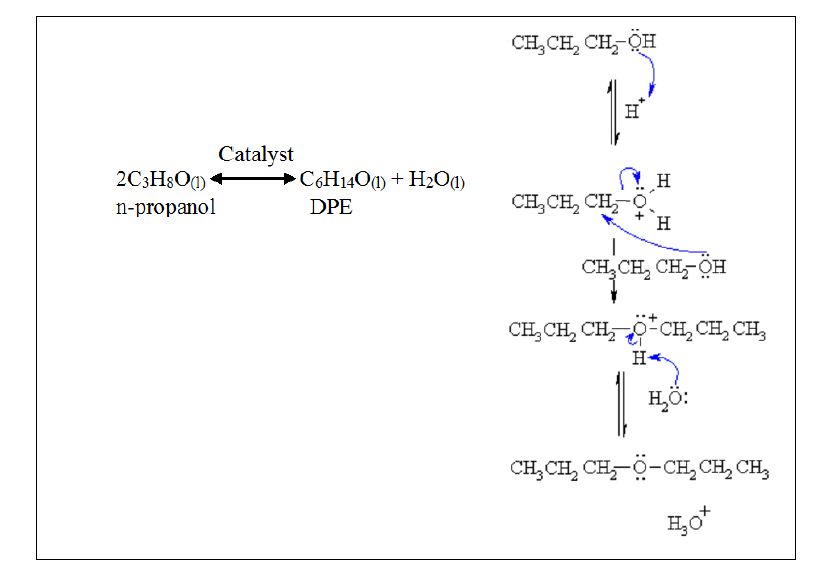
Due to the advances of biodiesel industry, a glut of glycerol has been resulted and there is a need for finding alternative uses for methanol contaminated glycerol. Due to the chemical composition of glycerol and methanol, dehydration and rearrangement could result in deoxygenated products that could be used as fuel additives. However, how methanol-containing glycerol could be converted into dehydrated products have not been widely investigated. This work attempts elucidating reaction conditions and performance parameters of an analogous system using methanol and n-propanol as the reactants while using catalysts that have been proven to be effective for transesterification.
Ethers consist of an oxygen atom bonded to two alkyl or aryl groups, or one alkyl and one aryl group [1]. Ethers have properties that might be beneficial as fuel additives to biodiesel and other biofuels. Boiling point is relatively low for ethers compared to their alcohol analogs which makes ethers more safe. Also, ethers have good solubility in hydrophilic and hydrophobic solvents making them compatible with esters such as biodiesel [2].
Ethers can be synthesized through several methods. The most common ones are bimolecular dehydration and Williamson method [1,3]. In dehydration reaction, alcohol is treated with a strong acid under dehydrating conditions. The general equation for dehydration reaction would be as follows [4]:
| $\text{ROH + R }\!\!'\!\!\text{ OH}\leftrightarrow \text{ROR }\!\!'\!\!\text{ + ROR+ R }\!\!'\!\!\text{ OR }\!\!'\!\!\text{ }+{{\text{H}}_{\text{2}}}\text{O}$ | (1) |
More specifically, above reaction may proceed according to following elementary steps:
ROH ↔ ROR
R'OH ↔ R'OR'
ROH + R'OH ↔ ROR'
In this work, we will use bimolecular dehydration to produce di-propyl ether (DPE) and methyl propyl ether (MPE) from methanol and n-propanol. It is anticipated that when a mixture of two alcohols was reacted through etherification, the result would be a mixture of three types of ethers as given in Scheme (1). In this case, three products, i.e., DPE, MPE, and dimethyl ether are anticipated.
Ethers that could result from methanol-contaminated glycerol dehydration, i.e., DPE and MPE have been of interest in chemical and pharmaceutical industry. DPE, a common oxygenated hydrocarbon, is used widely in industry as a solvent [5,6]. MPE is an isomer of diethyl ether, and has found many applications such as analgesic reagent [7], anesthetic [8] in the medical industry and a solvent (reference) and fuel additive [9] in the chemical industry.
DPE can be prepared from reacting 1-propanol by dehydration [10]. It has been predicted that DPE can be obtained through dehydration reaction from an already separated mixture of DPE and n-propanol using sulfuric acid as a catalyst. The mixture would be separated using extractive distillation and pressure-swing distillation [10]. Another general example is producing diethyl ether from ethanol dehydration. This work was done in the range of temperature of 140-250 °C with three different catalysts [11]. 1-Butanol has also been used to produce di-butyl ether; in that work, acidic ion-exchange resin Amberlyst 36 was used as the catalyst [12].
MPE has been produced as fraction of biocrude produced using Hydrothermal Upgrading (HTU) process by thermochemical conversion of biomass. This process requires high temperature and pressure, around 350 °C and 180 bar respectively [13]. However, MPE comprised of only 2.5% of the bio-crude produced. Other work done on methanol-isobutanol, ethanol-isobutanol and, ethanol etherification [11] also followed dehydration principle [14]. Above examples show that dehydration is a feasible method to produce ethers using alcohols.
Work to date on dehydration reveals that the yields are still low. Also, the high temperatures and pressures required make the process energy intensive and less economical. Additionally, there is little work has been done on strategies to increase the yield, conversion, and selectivity of ethers produced from common alcohols. This present work is geared toward identifying key parameters that impact alcohol conversion and ether yields and selectivity using methanol and 1-propanol with the intention of identify the best combination of parameter that would cost effectively produce dehydrated ethers. The novelty of this work is the study of the impact of three different catalysts that have distinct properties, i.e., sulfuric acid (a homogeneous duel proton donor), Amberlyst 36 (a heterogeneous strongly acidic cation exchanger) and titanium isopropoxide (a condensable base in the isopropoxide form that becomes acidic once condensed to TiO2).
Initial studies were conducted to establish catalysts and conditions that promote n-propanol etherification to form di-propyl ether. The rationale was that these variables would be a good starting point for MPE synthesis using methanol and n-propanol.
Studies with n-propanol were conducted in the presence of 1% (w/w) sulfuric acid, Amberlyst-36, and titanium isopropoxide. The reaction temperatures ranged from 100-160 °C at 20 °C intervals with a reaction time of 4 hrs.
Etherification studies on n-propanol methanol mix was conducted with select catalysts identified based on proposal etherification studies under the same conditions. The only exception was that in this case, 1% and 5% (w/w) of catalyst concentrations were tested. In this case, three products, i.e., DPE, MPE, and dimethyl ether (DME) are anticipated. However, we will report DPE and MPE data as a result of DME being a gaseous product and is of less use as a liquid fuel additive.
Response Measurements:
Primary variables that were calculated include substrate conversion, product yield and selectivity. For the case with n-propanol, aforementioned variables were calculated as follows:
Yield of DPE = $\frac{{{\text{n}}_{\left( \text{DPE} \right)}}}{\text{n}_{\left( \text{n}-\text{propanol} \right)}^{\text{o}}}$
Selectivity toward DPE = $\frac{{{\text{n}}_{\left( \text{DPE} \right)}}}{\text{n}_{\left( \text{n}-\text{propanol} \right)}^{0}+{{\text{n}}_{\left( \text{n}-\text{propanol} \right)}}}$
Propanol conversion = $\frac{\text{n}_{\left( \text{n}-\text{propanol} \right)}^{0}-{{\text{n}}_{\left( \text{n}-\text{propanol} \right)}}}{\text{n}_{\left( \text{n}-\text{propanol} \right)}^{0}}$
Where: n = final number of moles; no = initial number of moles.
When a methanol and propanol mixture was used, the variables were calculated as follows:
Ether yields:
Yield of DPE = $\frac{{{\text{n}}_{\left( \text{DPE} \right)}}}{\text{n}_{\left( \text{n}-\text{propanol} \right)}^{0}}$
Yield of MPE = $~\frac{{{n}_{\left( MPE \right)}}}{n_{\left( n-propanol \right)~}^{0}+~n_{\left( methanol \right)~}^{0}}$
Ether selectivity:
Selectivity toward DPE = $\frac{{{\text{n}}_{\left( \text{DPE} \right)}}}{{{\text{n}}_{\left( \text{DPE} \right)}}+{{\text{n}}_{\left( \text{MPE} \right)}}}$
Selectivity towards MPE = $\frac{{{\text{n}}_{\left( \text{MPE} \right)}}}{{{\text{n}}_{\left( \text{DPE} \right)}}+{{\text{n}}_{\left( \text{MPE} \right)}}}$
Substrate conversion:
Propanol conversion = $\frac{\text{n}_{\left( \text{n}-\text{propanol} \right)}^{0}-{{\text{n}}_{\left( \text{n}-\text{propanol} \right)}}}{\text{n}_{\left( \text{n}-\text{propanol} \right)}^{0}}$
Methanol Conversion = $\frac{\text{n}_{\left( \text{methanol} \right)}^{0}-{{\text{n}}_{\left( \text{methanol} \right)}}}{\text{n}_{\left( \text{methanol} \right)}^{0}}$
Where: n = number of moles; ${{n}^{0}}$ = initial number of moles.
Three catalysts were used: Sulfuric acid was obtained from J.T. Baker with an assay of 95.9%; Amberlyst 36 was provided from Sigma Aldrich with a water content of (51-57) % and a total pore volume of 0.2 mL/g, particle size 0.6-0.85 mm, surface area 33 m2/g, >1.95 eq/L exchange capacity; Titanium isopropoxide (colorless to yellow liquid with a density of 0.96 g/mL at 20 °C, boiling point of 232 °C, and molecular weight of 284.22 g/mol) was provided by Alfa Aesar company with >97% purity. All the catalysts were used at 5% w/w concentration.
Other reactants used were n-propanol and di-propyl ether and were obtained from Sigma Aldrich with a purity >99%.
Methanol (99.8% assay with a maximum of 0.1% water content) was obtained from VWR. Other reactants, n-propanol (99.7% assay), di-propyl ether (>99% assay) and MPE (97% assay), were obtained from Sigma Aldrich. Methyl-propyl-ether (Methoxypropane) and Di-propyl-ether were used as standards.
The reactions were carried out under stirring in a 25-mL high pressure vessel (Buchiglas USA) rated for 100 bar at 200 °C with temperature/pressure readout (it should be noted that the pressure in the vessel varied based on reaction temperature, and type and concentration of catalyst. The pressure in general ranged between (5-30) bar for Amberlyst 36, and between (10-40) bar for sulfuric catalyzed reaction). Weight measurements were carried out in a scale (PA 120) (with 0.001 mg sensitivity). Quantitative product analyses were carried out in a Gas Chromatograph (Agilent Technologies Model 6850) and Liquid Chromatograph (Agilent Technologies Model 1120 Compact LC).
The reaction between two n-propanol molecules in the presence of acid catalysts is expected to proceed according to Scheme (2) as follows:
The reaction is exothermic; however, energy input is necessary to overcome kinetic limitations. Therefore, the reaction progresses only with adequate energy input.
In terms of the methods, initially, the empty weight of a 32-mL vial was recorded. Then, reactants were added into the vial in a stepwise manner starting with 4 mL of n-propanol and 5% w/w of catalyst (note: equal catalyst weights were used since catalytic activity comparisons in biodiesel production generally uses mass basis rather than proton concentrations). The weight of the vial after addition of the chemical(s) was recorded. When the catalysts were solid, the weight of the catalysts was recorded separately. The chemicals and the magnetic stir-bar were added to the reactor securely sealed and placed in a stir hotplate (with stirring speed 700-800 rpm) for the reactions to progress. Once the reaction was over, the products were swiftly transferred to collection vials and sent for further analyses (GC, LC or weight/volume measurement) as needed.
The reaction between methanol and propanol progresses according to Scheme (3) as follows:
|
$\left. \right\}$
|
(3) |
The procedure for methanol and n-propanol etherification was similar to above except that the amounts of the reactants were different, i.e., 2 mL of methanol and 2 mL of n-propanol. The catalysts used in this case were sulfuric acid and Amberlyst-36 at 1% and 5% w/w concentrations.
The calibration standard consisted of methanol, n-propanol, methoxypropane, and di-propyl ether. Standard curves were developed by mixing above chemicals in predetermined ratios. When necessary, an internal standard (ISTD) was also used. An ISTD was used to account for any errors associated with sample handling. This was done by incorporating a correction factor as a result of internal standard analyses.
The top phase of each sample was analyzed via the GC. Each sample consisted of 1 mL total volume. The GC method consisted of following parameters:
· Column Information: Model (J&W 122-703E), Capillary Column with dimensions (30 m × 250 μm × 0.25 μm)
· Inlet type (EPC split-splitless inlet), and temperature is 220 °C
· Outlet Temperature: 240 °C
· Oven Temperature: 250 °C
· Gas Carrier and flow rate: Helium, 1.0 mL/min
· Detector: Flame Ionized Detector
The bottom aqueous phase was analyzed using HPLC. After separation from the top phase, the bottom phase was neutralized and filtered (to remove any solid material) before injecting to the HPLC. Neutralization was performed using NaOH for acid catalyzed reactions. During neutralization, the neutralizing solution (NaOH) was added dropwise while stirring and monitoring pH. The sample was considered neutral when the pH reached 7.0 ± 0.1. Sample filtration was done sequentially using 0.8 μm, 0.45 μm, and 0.25 μm syringe filters respectively. Then, samples were diluted as appropriate to be compatible with HPLC column and injected manually to HPLC. The HPLC method consisted of following parameters:
· Column Information: Supelcogel 610H (30 cm × 7.8 mm).
· Flow rate: 0.5 mL/min.
· Mobile phase: 0.1% phosphoric acid.
The n-propanol etherification was done as a Completely Randomized Design (CRD) that consisted of 12 units whereas methanol and propanol etherification was done as a full factorial design. In this design, all the variables and levels were randomized with three replicates using JMP software (SAS Institute). Results were obtained, organized, and analyzed using JMP software as well.
Substrate conversion and DPE yields & selectivity information is depicted in Figure 1. According to Figure 1A, the conversion clearly increased with increasing temperature for all the catalysts tested. Sulfuric acid clearly resulted in higher substrate conversions as compared to the other two catalysts for each temperature. Interestingly, at 140 and 160 °C, sulfuric acid resulted in complete conversion of di-propyl ether. Titanium isopropoxide performed the least with the highest conversion being only ~20% at 160 °C.
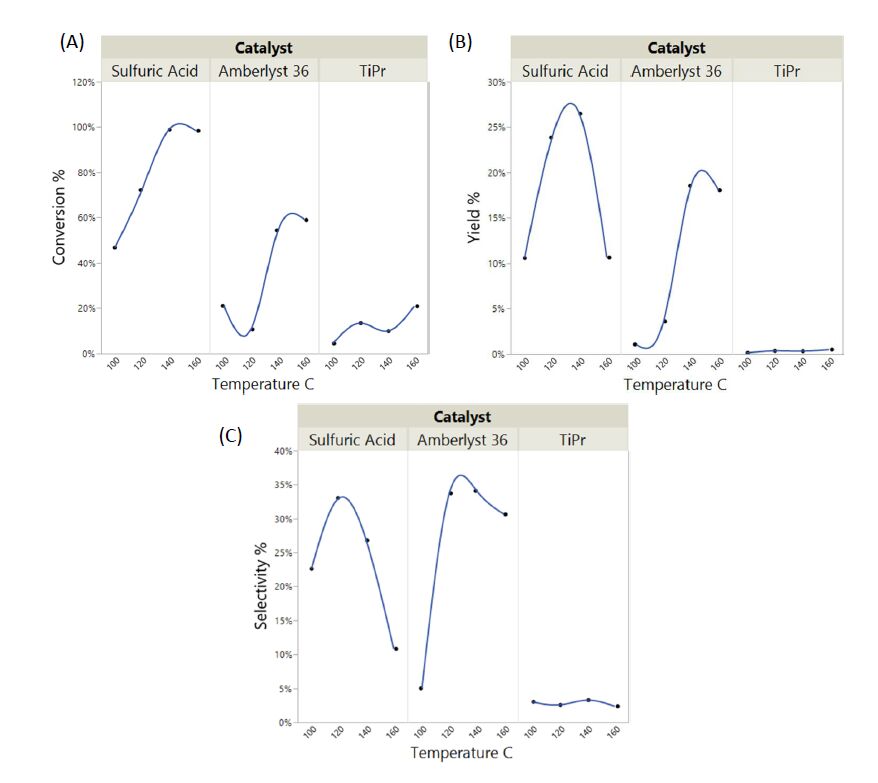 Figure 1. (A) n-propanol conversion; (B) di-propyl ether yield; and (C) selectivity toward di-propyl ether as a function of temperature and type of catalyst.
Figure 1. (A) n-propanol conversion; (B) di-propyl ether yield; and (C) selectivity toward di-propyl ether as a function of temperature and type of catalyst.DPE yields followed a similar trend to propanol conversion (Figure 1B). Again, sulfuric acid resulted in the highest DPE yields at 140 °C. It was interesting to note that the yields dropped at 160 °C, likely due to product disintegration. DPE yields increased with increasing temperature when Amberlyst-36 was used as the catalyst; however, the yields were lower as compared to when sulfuric acid was used. No significant ester yields were observed with titanium isopropoxide.
It should be noted that based on the selectivity equation used, the maximum selectivity obtainable in this situation is 50%. Interestingly, Amberlyst-36 resulted in the best selectivity toward DPE from the three catalysts tested. Sulfuric acid performed similar to Amberlyst-36 but with slightly lower overall selectivity values. It was evident in both cases, i.e., sulfuric acid and Amberlyst-36, that there was an optimum temperature that renders highest product selectivity which was ~120 °C for both catalysts. Again, titanium isopropoxide did not show any appreciable selectivity toward etherification.
The impact of temperature and type of catalyst on propanol conversion, DPE yield and the selectivity toward DPE are given in Table 1.
ANOVA indicates that temperature alone doesn't impact the conversion, yield or selectivity; however, the type of catalyst has a significant impact on all of the above responses. How the temperature and type of catalyst impacted propanol conversion, DPE yield and the selectivity toward DPE are depicted in Figure 1A, B and C respectively.
| Factor | Conversion | Yield of DPE | Selectivity towards DPE |
| Temperature °C | P = 0.0502 | P = 0.3526 | P = 0.3785 |
| Type of Catalyst | P = 0.0012 | P = 0.0312 | P = 0.0249 |
| P < 0.05 indicates significance | |||
 DownLoad: CSV
DownLoad: CSVSubstrate Conversion:The impact of catalyst type and concentration and temperature on n-propanol and methanol conversion is depicted in Figure 2.
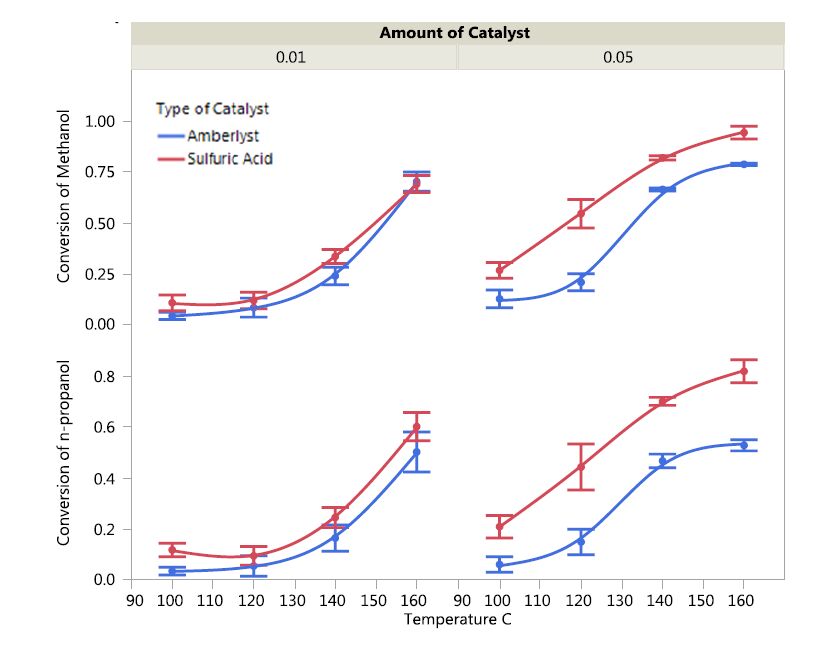 Figure 2. The impact of catalyst type, catalyst concentration, and temperature on conversion of n-propanol and methanol.
Figure 2. The impact of catalyst type, catalyst concentration, and temperature on conversion of n-propanol and methanol.It can be noted that sulfuric acid resulted in higher conversions of both n-propanol and methanol as compared to Amberlyst-36 even with the same concentration. This superiority in activity was evident at higher catalyst concentrations. This is likely as a result of the higher proton content in the case of sulfuric acid (twice as much protons) as compared to Amberlyst 36 per unit mass basis. The higher performance of sulfuric acid could also be attributed to the catalyst being homogeneous. Homogeneous catalysts generally perform better than heterogeneous catalysts due to much favorable mass transport characteristics in reactions [16,17,18]. Clearly, higher temperatures favored conversion of both substrates regardless of the type of catalyst; however, in a non-linear fashion. The better performance at higher temperature could be attributed to better reaction kinetics [19]. It was evident that methanol conversion was higher as compared to n-propanol and this could be attributed to more favorable steric of methanol (being the smaller of the two). Also, it should be noted that since methanol is initially in excess in the reaction medium, the fact that methanol conversion is higher than that of n-propanol implies that most of methanol may have converted in to byproducts such as dimethyl ether.
The yields of DPE and MPE as a function of catalyst type and amount for different temperatures are depicted in Figure 3.
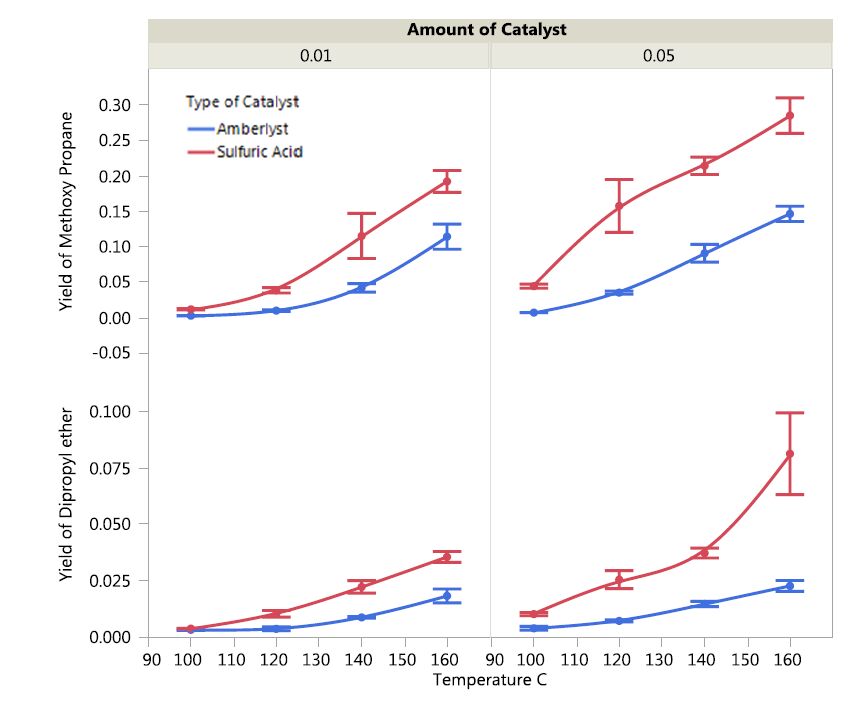 Figure 3. Impact of type and amount of catalysts on ether yield as a function of temperature.
Figure 3. Impact of type and amount of catalysts on ether yield as a function of temperature.Overall, sulfuric acid resulted in higher DPE yields as compared to Amberlyst 36 over the temperature range tested. DPE yields increased with increasing temperature for both catalysts. The DPE yield was 7.5% with 5% sulfuric acid catalyst at 160 °C. Although the DPE yields increased as temperature increased with both catalysts, Amberlyst lagged sulfuric acid in all instances. In terms of MPE yields, sulfuric acid resulted in better overall yields at all temperatures and catalyst concentrations as compared to Amberlyst 36. The yields tended to increase linearly with temperature for both catalysts.
The selectivity of the two catalysts, i.e., sulfuric acid and Amberlyst 36 toward production of and di-propyl ether are depicted in Figure 4.
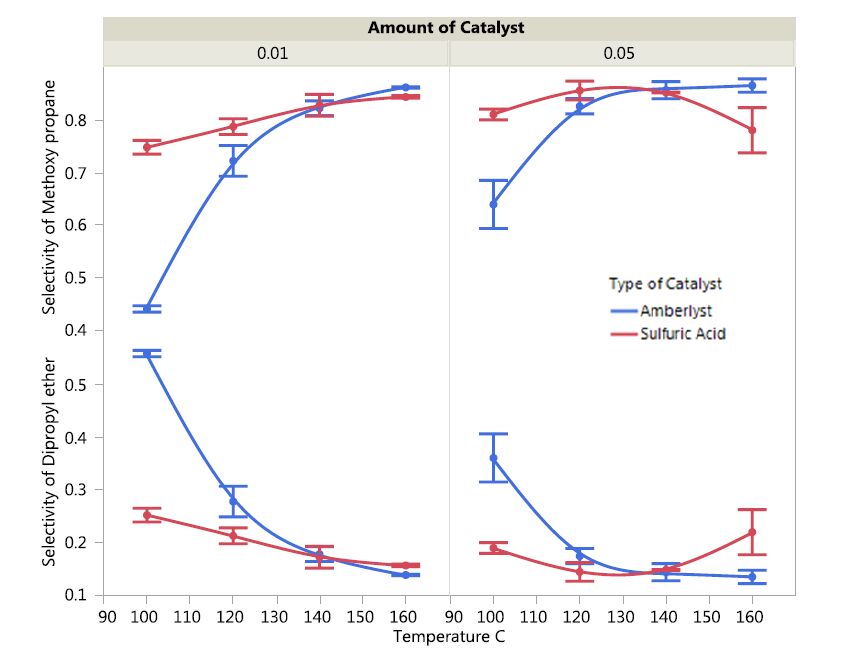 Figure 4. The impact of catalyst type and concentration on ether selectivity as a function of temperature.
Figure 4. The impact of catalyst type and concentration on ether selectivity as a function of temperature.It was interesting to note that the selectivity behavior of the catalysts for di-propyl ether was the antithesis to that of methoxypropane. In general, higher temperatures favored selectivity of both catalysts toward methoxypropane. At low temperatures, sulfuric acid showed markedly higher selectivity toward methoxypropane production as compared to dimethyl ether. However, as temperatures increased, the selectivity of Amberlyst 36 reached the same levels as sulfuric acid (at temperatures around 130 °C and even surpassed the values of sulfuric acid at higher temperatures). On the other hand, the selectivity of both catalysts toward di-propyl ether production was highest at low temperature and steadily declined as temperature increased. Amberlyst 36 was comparatively more selective toward di-propyl ether production as compared to production of methoxypropane. Also, both catalysts were comparatively more selective for methoxypropane production as opposed to production of di-propyl ether.
The impact of catalyst type, concentration and temperature on n-propanol and methanol conversion and yield and selectivity toward the products, di-propyl ether (DPE) and methyl-propyl ether (MPE) are presented in Table 2.
| Responses | ||||||
| Factor | Conversion of Methanol | Conversion of n-propanol | Yield of DPE | Yield of MPE | Selectivity towards DPE | Selectivity toward MPE |
| Temperature, °C | P =<0.0001 | P =<0.0001 | P =<0.0001 | P = 0.595 | P =<0.0001 | P =<0.0001 |
| Type of Catalyst | P = 0.0003 | P =<0.0001 | P =<0.0001 | P = 0.9 | P = 0.0181 | P = 0.0181 |
| Catalyst Concentration | P =<0.0001 | P =<0.0001 | P = 0.0010 | P = 0.0295 | P = 0.0295 | P = 0.0295 |
| Catalyst Concentration *Type of Catalyst | P = 0.0181 | P = 0.0089 | P = 0.0176 | P = 0.8815 | P = 0.2027 | P = 0.2027 |
| *P: Probability value for significance. P < 0.05 indicates significance. | ||||||
DownLoad: CSVIt can be seen that with the exception of catalyst concentration vs type interaction on product selectivity, all other factors significantly impacted the responses. This implies that applying different combinations of levels of the variables matter when it comes to substrate conversion, product yield and to a great extent, selectivity toward ethers.
In general, sulfuric acid resulted in comparatively higher conversions and ether yields. However, Amberlyst 36 gave comparable values of MPE yields and selectivity to sulfuric acid, especially when considering MPE. The high activity of sulfuric acid is likely as a result of the catalyst having a higher number of protons per mass basis. Sulfuric acid has twice number of protons as compared to Amberlyst 36. Also, it is possible that sulfuric acid being homogeneous would explain the higher activity (as opposed to Amberlyst being a heterogeneous catalyst) [20]. Homogeneous catalysts in general have a mass transport advantage in comparison to heterogeneous catalysts [18].
The higher selectivity of Amberlyst 36 for DPE production and surpassing the selectivity toward MPE at higher temperatures is interesting. It has been revealed during recent experiments that Amberlyst 36 has swelling capability [21]. This swelling capability has been associated with Amberlyst's ability to absorb water and polar components [22]. Karl-Fisher titration has revealed that the bottom product phase contains about 67% water. The ability of Amberlyst 36 to help remove the produced more efficiently than sulfuric acid may at least partially explain why Amberlyst 36 performed better under some conditions.
A mass balance was conducted for the reaction that consisted with 1:1 methanol to n-propanol volume ratio (i.e., 0.128 mol (4.08 g) of methanol, 0.064 mol (3.83 g) of propanol) with 5% sulfuric acid for 4 h. The products on average were 0.0215 mol (1.59 g) of MPE (~20% w/w) and 0.0048 mol (0.49 g) of DPE (~6%). The rest 5.83 g (~74%) were distributed among bottom phase (water and unconverted alcohol), and gaseous products.
Based on the above experiments, it is clear that nearly all the variables tested, i.e., catalyst type & concentration and reaction temperature, had some impact on the substrate conversion and product yield & selectivity. All the variables taken together, higher temperatures (i.e., 160 °C) seems to favor higher substrate conversion. However, if the objective is to promote coupling of larger alcohols, somewhat milder temperatures seem to favor higher product yield and selectivity. Of the two catalysts, sulfuric acid consistently produced higher conversion and DPE yield and selectivity. However, amberlyst 36 gave higher product yields, and selectivity for MPE.
Etherification experiments of propanol to di-propyl ether in the presence of sulfuric acid, Amberlyst 36 and titanium isopropoxide between temperatures 100-160 °Cindicated that the type of catalyst had a significant impact on substrate conversion, product yield, and product selectivity. Of the three catalysts, sulfuric acid and Amberlyst 36 performed the best while titanium isopropoxide did not show any significant activity toward etherification. Propanol conversion, and ether yields & selectivity increased with increasing temperature but peaked around 140 °C. Further studies with sulfuric acid and Amberlyst-36 at 1 and 5% (w/w) concentrations with methanol and n-propanol substrates revealed that higher catalyst concentrations and higher temperatures favored substrate conversion. Of the two catalysts, sulfuric acid resulted in better substrate conversion, and ether yields. Regardless of the catalyst, higher temperatures favored ether yields. It was interesting to note that the selectivity of the two catalysts toward di-propyl ether under increasing temperatures was the antithesis to methoxypropane. A significant finding of this study is that when two different sized alcohols are present, preferential coupling, i.e., if the larger alcohols are allowed to couple together or larger ones with small ones, could be controlled by varying the type of catalyst (sulfuric acid or Amberlyst 36). These results pave way to finding the correct catalyst type, concentration and conditions for allowing coupling more complex alcohols such as glycerol and methanol which will allow use of byproducts from reactions such as transesterification to useful chemicals and fuels.
The financial support provided by Texas AgriLife Research and the Higher Committee for Education Development in Iraq (HCED) is gratefully acknowledged.
The authors declare there is no conflict of interest.
| [1] |
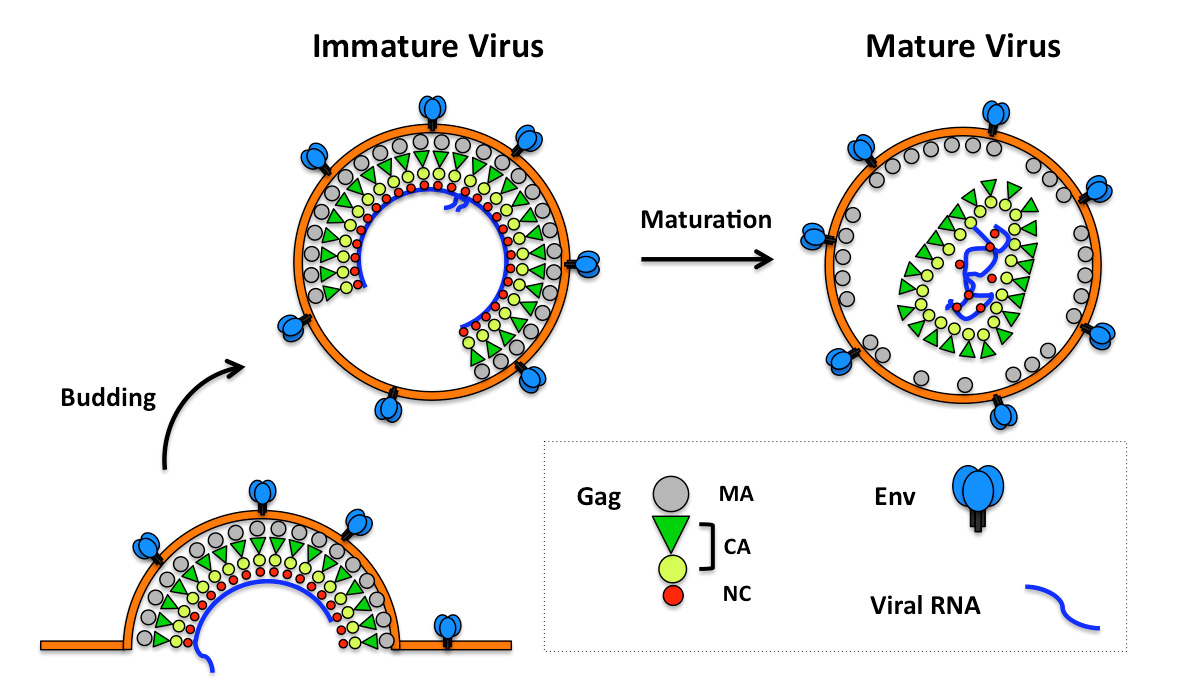
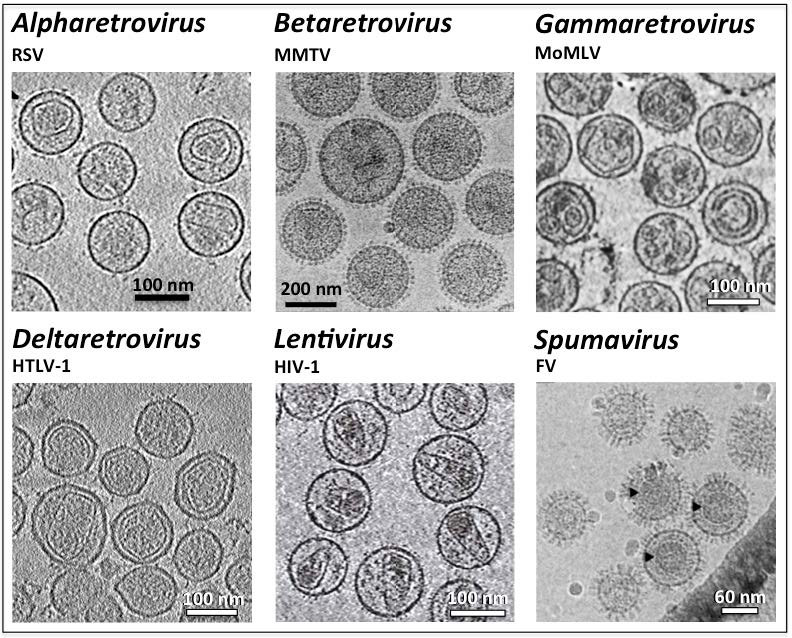
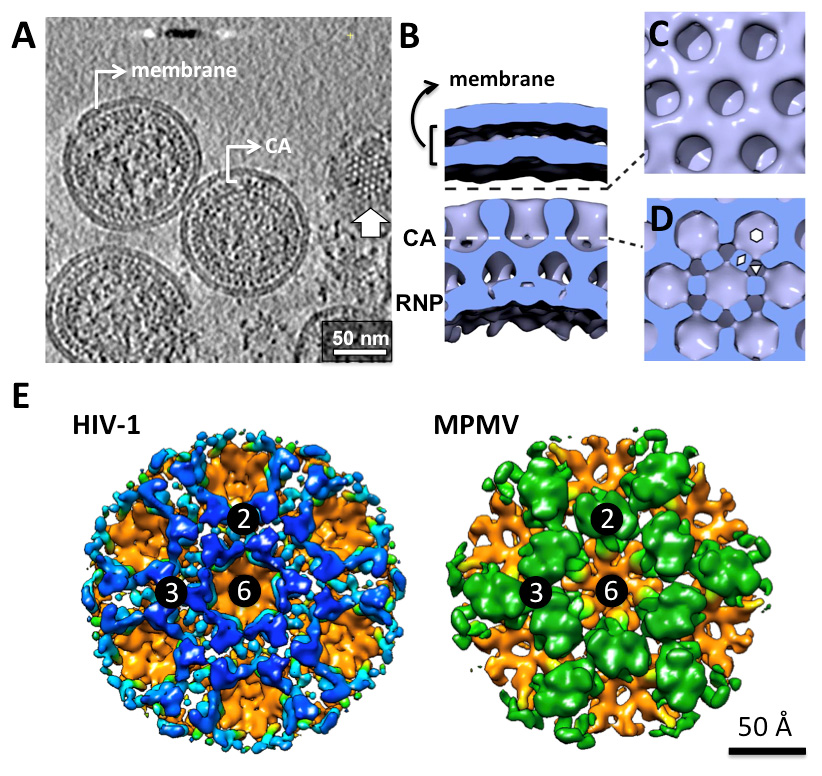
Ganser-Pornillos BK, Yeager M, Pornillos O (2012) Assembly and architecture of HIV. Adv Exp Med Biol 726: 441-465. doi: 10.1007/978-1-4614-0980-9_20

|
| [2] | Sundquist WI, Krausslich HG (2012) HIV-1 assembly, budding, and maturation. Cold Spring Harb Perspect Med 2: a006924. |
| [3] | Borggren M, Jansson M (2015) The Evolution of HIV-1 Interactions with Coreceptors and Mannose C-Type Lectin Receptors. Prog Mol Biol Transl Sci 129C: 109-140. |
| [4] |
Miyauchi K, Kim Y, Latinovic O, et al. (2009) HIV enters cells via endocytosis and dynamin-dependent fusion with endosomes. Cell 137: 433-444. doi: 10.1016/j.cell.2009.02.046
|
| [5] |
Moulard M, Decroly E (2000) Maturation of HIV envelope glycoprotein precursors by cellular endoproteases. Biochim Biophys Acta 1469: 121-132. doi: 10.1016/S0304-4157(00)00014-9
|
| [6] |
Low JT, Garcia-Miranda P, Mouzakis KD, et al. (2014) Structure and dynamics of the HIV-1 frameshift element RNA. Biochemistry 53: 4282-4291. doi: 10.1021/bi5004926
|
| [7] |
Stephenson JD, Li H, Kenyon JC, et al. (2013) Three-dimensional RNA structure of the major HIV-1 packaging signal region. Structure 21: 951-962. doi: 10.1016/j.str.2013.04.008
|
| [8] |
Watts JM, Dang KK, Gorelick RJ, et al. (2009) Architecture and secondary structure of an entire HIV-1 RNA genome. Nature 460: 711-716. doi: 10.1038/nature08237
|
| [9] | Swanstrom R, Wills JW (1997) Synthesis, Assembly, and Processing of Viral Proteins. In: Coffin JM HS, Varmus HE, editor. Retroviruses. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press. |
| [10] |
Checkley MA, Luttge BG, Freed EO (2011) HIV-1 envelope glycoprotein biosynthesis, trafficking, and incorporation. J Mol Biol 410: 582-608. doi: 10.1016/j.jmb.2011.04.042
|
| [11] |
Postler TS, Desrosiers RC (2013) The tale of the long tail: the cytoplasmic domain of HIV-1 gp41. J Virol 87: 2-15. doi: 10.1128/JVI.02053-12
|
| [12] |
Bharat TA, Davey NE, Ulbrich P, et al. (2012) Structure of the immature retroviral capsid at 8 A resolution by cryo-electron microscopy. Nature 487: 385-389. doi: 10.1038/nature11169
|
| [13] | Maldonado JO, Martin JL, Mueller JD, et al. (2014) New insights into retroviral Gag-Gag and Gag-membrane interactions. Front Microbiol 5: 302. |
| [14] | Sonnino S, Prinetti A (2013) Membrane domains and the “lipid raft” concept. Curr Med Chem 20: 4-21. |
| [15] |
Levin JG, Mitra M, Mascarenhas A, et al. (2010) Role of HIV-1 nucleocapsid protein in HIV-1 reverse transcription. RNA Biol 7: 754-774. doi: 10.4161/rna.7.6.14115
|
| [16] |
Jiang J, Aiken C (2007) Maturation-dependent human immunodeficiency virus type 1 particle fusion requires a carboxyl-terminal region of the gp41 cytoplasmic tail. J Virol 81: 9999-10008. doi: 10.1128/JVI.00592-07
|
| [17] |
Wyma DJ, Jiang J, Shi J, et al. (2004) Coupling of human immunodeficiency virus type 1 fusion to virion maturation: a novel role of the gp41 cytoplasmic tail. J Virol 78: 3429-3435. doi: 10.1128/JVI.78.7.3429-3435.2004
|
| [18] | Aiken C, Zhang P (2013) HIV-1 Maturation. In: Freed EO, editor. Advances in HIV-1 Assembly and Release: Springer. |
| [19] |
Forshey BM, von Schwedler U, Sundquist WI, et al. (2002) Formation of a human immunodeficiency virus type 1 core of optimal stability is crucial for viral replication. J Virol 76: 5667-5677. doi: 10.1128/JVI.76.11.5667-5677.2002
|
| [20] |
Ganser-Pornillos BK, Yeager M, Sundquist WI (2008) The structural biology of HIV assembly. Curr Opin Struct Biol 18: 203-217. doi: 10.1016/j.sbi.2008.02.001
|
| [21] |
Bush DL, Vogt VM (2014) In Vitro Assembly of Retroviruses. Annual Reviews in Virology 1: 561-580. doi: 10.1146/annurev-virology-031413-085427
|
| [22] |
Briggs JA (2013) Structural biology in situ--the potential of subtomogram averaging. Curr Opin Struct Biol 23: 261-267. doi: 10.1016/j.sbi.2013.02.003
|
| [23] |
Merk A, Subramaniam S (2013) HIV-1 envelope glycoprotein structure. Curr Opin Struct Biol 23: 268-276. doi: 10.1016/j.sbi.2013.03.007
|
| [24] |
Briggs JA, Simon MN, Gross I, et al. (2004) The stoichiometry of Gag protein in HIV-1. Nat Struct Mol Biol 11: 672-675. doi: 10.1038/nsmb785
|
| [25] |
Butan C, Winkler DC, Heymann JB, et al. (2008) RSV capsid polymorphism correlates with polymerization efficiency and envelope glycoprotein content: implications that nucleation controls morphogenesis. J Mol Biol 376: 1168-1181. doi: 10.1016/j.jmb.2007.12.003
|
| [26] |
Fuller SD, Wilk T, Gowen BE, et al. (1997) Cryo-electron microscopy reveals ordered domains in the immature HIV-1 particle. Curr Biol 7: 729-738. doi: 10.1016/S0960-9822(06)00331-9
|
| [27] |
Kingston RL, Olson NH, Vogt VM (2001) The organization of mature Rous sarcoma virus as studied by cryoelectron microscopy. J Struct Biol 136: 67-80. doi: 10.1006/jsbi.2001.4423
|
| [28] |
Yeager M, Wilson-Kubalek EM, Weiner SG, et al. (1998) Supramolecular organization of immature and mature murine leukemia virus revealed by electron cryo-microscopy: implications for retroviral assembly mechanisms. Proc Natl Acad Sci U S A 95: 7299-7304. doi: 10.1073/pnas.95.13.7299
|
| [29] |
Carlson LA, Briggs JA, Glass B, et al. (2008) Three-dimensional analysis of budding sites and released virus suggests a revised model for HIV-1 morphogenesis. Cell Host Microbe 4: 592-599. doi: 10.1016/j.chom.2008.10.013
|
| [30] |
Briggs JA, Johnson MC, Simon MN, et al. (2006) Cryo-electron microscopy reveals conserved and divergent features of gag packing in immature particles of Rous sarcoma virus and human immunodeficiency virus. J Mol Biol 355: 157-168. doi: 10.1016/j.jmb.2005.10.025
|
| [31] |
Briggs JA, Riches JD, Glass B, et al. (2009) Structure and assembly of immature HIV. Proc Natl Acad Sci U S A 106: 11090-11095. doi: 10.1073/pnas.0903535106
|
| [32] |
Keller PW, Huang RK, England MR, et al. (2013) A two-pronged structural analysis of retroviral maturation indicates that core formation proceeds by a disassembly-reassembly pathway rather than a displacive transition. J Virol 87: 13655-13664. doi: 10.1128/JVI.01408-13
|
| [33] |
Wright ER, Schooler JB, Ding HJ, et al. (2007) Electron cryotomography of immature HIV-1 virions reveals the structure of the CA and SP1 Gag shells. EMBO J 26: 2218-2226. doi: 10.1038/sj.emboj.7601664
|
| [34] |
de Marco A, Davey NE, Ulbrich P, et al. (2010) Conserved and variable features of Gag structure and arrangement in immature retrovirus particles. J Virol 84: 11729-11736. doi: 10.1128/JVI.01423-10
|
| [35] |
Datta SA, Temeselew LG, Crist RM, et al. (2011) On the role of the SP1 domain in HIV-1 particle assembly: a molecular switch? J Virol 85: 4111-4121. doi: 10.1128/JVI.00006-11
|
| [36] |
Liang C, Hu J, Russell RS, et al. (2002) Characterization of a putative alpha-helix across the capsid-SP1 boundary that is critical for the multimerization of human immunodeficiency virus type 1 gag. J Virol 76: 11729-11737. doi: 10.1128/JVI.76.22.11729-11737.2002
|
| [37] |
Briggs JA, Wilk T, Welker R, et al. (2003) Structural organization of authentic, mature HIV-1 virions and cores. EMBO J 22: 1707-1715. doi: 10.1093/emboj/cdg143
|
| [38] |
Ganser BK, Li S, Klishko VY, et al. (1999) Assembly and analysis of conical models for the HIV-1 core. Science 283: 80-83. doi: 10.1126/science.283.5398.80
|
| [39] |
Heymann JB, Butan C, Winkler DC, et al. (2008) Irregular and Semi-Regular Polyhedral Models for Rous Sarcoma Virus Cores. Comput Math Methods Med 9: 197-210. doi: 10.1080/17486700802168106
|
| [40] |
Cao S, Maldonado JO, Grigsby IF, et al. (2015) Analysis of human T-cell leukemia virus type 1 particles using cryo-electron tomography. J Virol 89: 2430-2435. doi: 10.1128/JVI.02358-14
|
| [41] |
Li S, Hill CP, Sundquist WI, et al. (2000) Image reconstructions of helical assemblies of the HIV-1 CA protein. Nature 407: 409-413. doi: 10.1038/35030177
|
| [42] |
Thomas D, Schultz P, Steven AC, et al. (1994) Mass analysis of biological macromolecular complexes by STEM. Biol Cell 80: 181-192. doi: 10.1111/j.1768-322X.1994.tb00929.x
|
| [43] |
Fogarty KH, Zhang W, Grigsby IF, et al. (2011) New insights into HTLV-1 particle structure, assembly, and Gag-Gag interactions in living cells. Viruses 3: 770-793. doi: 10.3390/v3060770
|
| [44] |
Johnson J, Chen Y, Mueller JD (2010) Characterization of brightness and stoichiometry of bright particles by flow-fluorescence fluctuation spectroscopy. Biophys J 99: 3084-3092. doi: 10.1016/j.bpj.2010.08.057
|
| [45] |
Parker SD, Wall JS, Hunter E (2001) Analysis of Mason-Pfizer monkey virus Gag particles by scanning transmission electron microscopy. J Virol 75: 9543-9548. doi: 10.1128/JVI.75.19.9543-9548.2001
|
| [46] | Vogt VM, Simon MN (1999) Mass determination of rous sarcoma virus virions by scanning transmission electron microscopy. J Virol 73: 7050-7055. |
| [47] |
Yu F, Joshi SM, Ma YM, et al. (2001) Characterization of Rous sarcoma virus Gag particles assembled in vitro. J Virol 75: 2753-2764. doi: 10.1128/JVI.75.6.2753-2764.2001
|
| [48] |
Chen Y, Wu B, Musier-Forsyth K, et al. (2009) Fluorescence fluctuation spectroscopy on viral-like particles reveals variable gag stoichiometry. Biophys J 96: 1961-1969. doi: 10.1016/j.bpj.2008.10.067
|
| [49] |
Grigsby IF, Zhang W, Johnson JL, et al. (2010) Biophysical analysis of HTLV-1 particles reveals novel insights into particle morphology and Gag stochiometry. Retrovirology 7: 75. doi: 10.1186/1742-4690-7-75
|
| [50] |
Lanman J, Lam TT, Emmett MR, et al. (2004) Key interactions in HIV-1 maturation identified by hydrogen-deuterium exchange. Nat Struct Mol Biol 11: 676-677. doi: 10.1038/nsmb790
|
| [51] |
Briggs JA, Watson BE, Gowen BE, et al. (2004) Cryoelectron microscopy of mouse mammary tumor virus. J Virol 78: 2606-2608. doi: 10.1128/JVI.78.5.2606-2608.2004
|
| [52] |
Forster F, Medalia O, Zauberman N, et al. (2005) Retrovirus envelope protein complex structure in situ studied by cryo-electron tomography. Proc Natl Acad Sci U S A 102: 4729-4734. doi: 10.1073/pnas.0409178102
|
| [53] |
Briggs JA, Grunewald K, Glass B, et al. (2006) The mechanism of HIV-1 core assembly: insights from three-dimensional reconstructions of authentic virions. Structure 14: 15-20. doi: 10.1016/j.str.2005.09.010
|
| [54] |
Hamann MV, Mullers E, Reh J, et al. (2014) The cooperative function of arginine residues in the Prototype Foamy Virus Gag C-terminus mediates viral and cellular RNA encapsidation. Retrovirology 11: 87. doi: 10.1186/s12977-014-0087-7
|
| [55] |
Roux KH, Taylor KA (2007) AIDS virus envelope spike structure. Curr Opin Struct Biol 17: 244-252. doi: 10.1016/j.sbi.2007.03.008
|
| [56] |
Liu J, Bartesaghi A, Borgnia MJ, et al. (2008) Molecular architecture of native HIV-1 gp120 trimers. Nature 455: 109-113. doi: 10.1038/nature07159
|
| [57] |
Zanetti G, Briggs JA, Grunewald K, et al. (2006) Cryo-electron tomographic structure of an immunodeficiency virus envelope complex in situ. PLoS Pathog 2: e83. doi: 10.1371/journal.ppat.0020083
|
| [58] |
Harris A, Borgnia MJ, Shi D, et al. (2011) Trimeric HIV-1 glycoprotein gp140 immunogens and native HIV-1 envelope glycoproteins display the same closed and open quaternary molecular architectures. Proc Natl Acad Sci U S A 108: 11440-11445. doi: 10.1073/pnas.1101414108
|
| [59] |
Meyerson JR, Tran EE, Kuybeda O, et al. (2013) Molecular structures of trimeric HIV-1 Env in complex with small antibody derivatives. Proc Natl Acad Sci U S A 110: 513-518. doi: 10.1073/pnas.1214810110
|
| [60] |
Tran EE, Borgnia MJ, Kuybeda O, et al. (2012) Structural mechanism of trimeric HIV-1 envelope glycoprotein activation. PLoS Pathog 8: e1002797. doi: 10.1371/journal.ppat.1002797
|
| [61] |
White TA, Bartesaghi A, Borgnia MJ, et al. (2011) Three-dimensional structures of soluble CD4-bound states of trimeric simian immunodeficiency virus envelope glycoproteins determined by using cryo-electron tomography. J Virol 85: 12114-12123. doi: 10.1128/JVI.05297-11
|
| [62] |
White TA, Bartesaghi A, Borgnia MJ, et al. (2010) Molecular architectures of trimeric SIV and HIV-1 envelope glycoproteins on intact viruses: strain-dependent variation in quaternary structure. PLoS Pathog 6: e1001249. doi: 10.1371/journal.ppat.1001249
|
| [63] |
Loving R, Wu SR, Sjoberg M, et al. (2012) Maturation cleavage of the murine leukemia virus Env precursor separates the transmembrane subunits to prime it for receptor triggering. Proc Natl Acad Sci U S A 109: 7735-7740. doi: 10.1073/pnas.1118125109
|
| [64] |
Sjoberg M, Wu SR, Loving R, et al. (2014) Furin cleavage of the Moloney murine leukemia virus Env precursor reorganizes the spike structure. Proc Natl Acad Sci U S A 111: 6034-6039. doi: 10.1073/pnas.1317972111
|
| [65] |
Wu SR, Sjoberg M, Wallin M, et al. (2008) Turning of the receptor-binding domains opens up the murine leukaemia virus Env for membrane fusion. EMBO J 27: 2799-2808. doi: 10.1038/emboj.2008.187
|
| [66] |
Zhu P, Liu J, Bess J, Jr., et al. (2006) Distribution and three-dimensional structure of AIDS virus envelope spikes. Nature 441: 847-852. doi: 10.1038/nature04817
|
| [67] | Schur FK, Hagen WJ, Rumlova M, et al. (2015) Structure of the immature HIV-1 capsid in intact virus particles at 8.8 A resolution. . Nature 517: 505-508. |
| [68] |
Pettersen EF, Goddard TD, Huang CC, et al. (2004) UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem 25: 1605-1612. doi: 10.1002/jcc.20084
|
| [69] |
Zhao G, Perilla JR, Yufenyuy EL, et al. (2013) Mature HIV-1 capsid structure by cryo-electron microscopy and all-atom molecular dynamics. Nature 497: 643-646. doi: 10.1038/nature12162
|
| [70] |
Cardone G, Purdy JG, Cheng N, et al. (2009) Visualization of a missing link in retrovirus capsid assembly. Nature 457: 694-698. doi: 10.1038/nature07724
|
| [71] |
Hyun JK, Radjainia M, Kingston RL, et al. (2010) Proton-driven assembly of the Rous Sarcoma virus capsid protein results in the formation of icosahedral particles. J Biol Chem 285: 15056-15064. doi: 10.1074/jbc.M110.108209
|
| [72] |
Ganser-Pornillos BK, Cheng A, Yeager M (2007) Structure of full-length HIV-1 CA: a model for the mature capsid lattice. Cell 131: 70-79. doi: 10.1016/j.cell.2007.08.018
|
| [73] |
Bartesaghi A, Merk A, Borgnia MJ, et al. (2013) Prefusion structure of trimeric HIV-1 envelope glycoprotein determined by cryo-electron microscopy. Nat Struct Mol Biol 20: 1352-1357. doi: 10.1038/nsmb.2711
|
| [74] |
Lyumkis D, Julien JP, de Val N, et al. (2013) Cryo-EM structure of a fully glycosylated soluble cleaved HIV-1 envelope trimer. Science 342: 1484-1490. doi: 10.1126/science.1245627
|
| [75] |
Bartesaghi A, Lecumberry F, Sapiro G, et al. (2012) Protein secondary structure determination by constrained single-particle cryo-electron tomography. Structure 20: 2003-2013. doi: 10.1016/j.str.2012.10.016
|
| [76] | Schur FK, Hagen WJ, de Marco A, et al. (2013) Determination of protein structure at 8.5A resolution using cryo-electron tomography and sub-tomogram averaging. . J Struct Biol 184: 394-400. |
| [77] | Tang C, Ndassa Y, Summers MF (2002) Structure of the N-terminal 283-residue fragment of the immature HIV-1 Gag polyprotein. Nat Struct Biol 9: 537-543. |
| [78] |
Pornillos O, Ganser-Pornillos BK, Kelly BN, et al. (2009) X-ray structures of the hexameric building block of the HIV capsid. Cell 137: 1282-1292. doi: 10.1016/j.cell.2009.04.063
|
| [79] |
Macek P, Chmelik J, Krizova I, et al. (2009) NMR structure of the N-terminal domain of capsid protein from the mason-pfizer monkey virus. J Mol Biol 392: 100-114. doi: 10.1016/j.jmb.2009.06.029
|
| [80] |
Bharat TA, Castillo Menendez LR, Hagen WJ, et al. (2014) Cryo-electron microscopy of tubular arrays of HIV-1 Gag resolves structures essential for immature virus assembly. Proc Natl Acad Sci U S A 111: 8233-8238. doi: 10.1073/pnas.1401455111
|
| [81] |
Mao Y, Wang L, Gu C, et al. (2012) Subunit organization of the membrane-bound HIV-1 envelope glycoprotein trimer. Nat Struct Mol Biol 19: 893-899. doi: 10.1038/nsmb.2351
|
| [82] |
Mao Y, Wang L, Gu C, et al. (2013) Molecular architecture of the uncleaved HIV-1 envelope glycoprotein trimer. Proc Natl Acad Sci U S A 110: 12438-12443. doi: 10.1073/pnas.1307382110
|
| [83] | Cohen J Structural biology. Is high-tech view of HIV too good to be true? Science 341: 443-444. |
| [84] | Henderson R Avoiding the pitfalls of single particle cryo-electron microscopy: Einstein from noise. Proc Natl Acad Sci U S A 110: 18037-18041. |
| [85] | Mao Y, Castillo-Menendez LR, Sodroski JG Reply to Subramaniam, van Heel, and Henderson: Validity of the cryo-electron microscopy structures of the HIV-1 envelope glycoprotein complex. Proc Natl Acad Sci U S A 110: E4178-4182. |
| [86] |
Frank GA, Narayan K, Bess JW, Jr., et al. (2015) Maturation of the HIV-1 core by a non-diffusional phase transition. Nat Commun 6: 5854. doi: 10.1038/ncomms6854
|
| [87] | van Heel M Finding trimeric HIV-1 envelope glycoproteins in random noise. Proc Natl Acad Sci U S A 110: E4175-4177. |
| [88] | Mammano F, Ohagen A, Hoglund S, et al. (1994) Role of the major homology region of human immunodeficiency virus type 1 in virion morphogenesis. J Virol 68: 4927-4936. |
| [89] |
von Schwedler UK, Stray KM, Garrus JE, et al. (2003) Functional surfaces of the human immunodeficiency virus type 1 capsid protein. J Virol 77: 5439-5450. doi: 10.1128/JVI.77.9.5439-5450.2003
|
| [90] |
Chu HH, Chang YF, Wang CT (2006) Mutations in the alpha-helix directly C-terminal to the major homology region of human immunodeficiency virus type 1 capsid protein disrupt Gag multimerization and markedly impair virus particle production. J Biomed Sci 13: 645-656. doi: 10.1007/s11373-006-9094-6
|
| [91] |
Gamble TR, Yoo S, Vajdos FF, et al. (1997) Structure of the carboxyl-terminal dimerization domain of the HIV-1 capsid protein. Science 278: 849-853. doi: 10.1126/science.278.5339.849
|
| [92] |
Ono A, Ablan SD, Lockett SJ, et al. (2004) Phosphatidylinositol (4,5) bisphosphate regulates HIV-1 Gag targeting to the plasma membrane. Proc Natl Acad Sci U S A 101: 14889-14894. doi: 10.1073/pnas.0405596101
|
| [93] |
Saad JS, Miller J, Tai J, et al. (2006) Structural basis for targeting HIV-1 Gag proteins to the plasma membrane for virus assembly. Proc Natl Acad Sci U S A 103: 11364-11369. doi: 10.1073/pnas.0602818103
|
| [94] |
Fogarty KH, Chen Y, Grigsby IF, et al. (2011) Characterization of cytoplasmic Gag-gag interactions by dual-color z-scan fluorescence fluctuation spectroscopy. Biophys J 100: 1587-1595. doi: 10.1016/j.bpj.2011.02.008
|
| [95] |
Lindwasser OW, Resh MD (2004) Human immunodeficiency virus type 1 Gag contains a dileucine-like motif that regulates association with multivesicular bodies. J Virol 78: 6013-6023. doi: 10.1128/JVI.78.11.6013-6023.2004
|
| [96] |
Muriaux D, Mirro J, Harvin D, et al. (2001) RNA is a structural element in retrovirus particles. Proc Natl Acad Sci U S A 98: 5246-5251. doi: 10.1073/pnas.091000398
|
| [97] |
Poole E, Strappe P, Mok HP, et al. (2005) HIV-1 Gag-RNA interaction occurs at a perinuclear/centrosomal site; analysis by confocal microscopy and FRET. Traffic 6: 741-755. doi: 10.1111/j.1600-0854.2005.00312.x
|
| [98] |
Bailey GD, Hyun JK, Mitra AK, et al. (2012) A structural model for the generation of continuous curvature on the surface of a retroviral capsid. J Mol Biol 417: 212-223. doi: 10.1016/j.jmb.2012.01.014
|
| [99] |
Mortuza GB, Haire LF, Stevens A, et al. (2004) High-resolution structure of a retroviral capsid hexameric amino-terminal domain. Nature 431: 481-485. doi: 10.1038/nature02915
|
| [100] |
Byeon IJ, Meng X, Jung J, et al. (2009) Structural convergence between Cryo-EM and NMR reveals intersubunit interactions critical for HIV-1 capsid function. Cell 139: 780-790. doi: 10.1016/j.cell.2009.10.010
|
| [101] |
Yang H, Ji X, Zhao G, et al. (2012) Structural insight into HIV-1 capsid recognition by rhesus TRIM5alpha. Proc Natl Acad Sci U S A 109: 18372-18377. doi: 10.1073/pnas.1210903109
|
| [102] |
Zhao G, Ke D, Vu T, et al. (2011) Rhesus TRIM5alpha disrupts the HIV-1 capsid at the inter-hexamer interfaces. PLoS Pathog 7: e1002009. doi: 10.1371/journal.ppat.1002009
|
| [103] | Gres AT, Kirby KA, KewalRamani VN, et al. (2015) X-ray crystal structures of native HIV-1 capsid protein reveal conformational variability. Science. |
| [104] |
Dalgleish AG, Beverley PC, Clapham PR, et al. (1984) The CD4 (T4) antigen is an essential component of the receptor for the AIDS retrovirus. Nature 312: 763-767. doi: 10.1038/312763a0
|
| [105] |
Trkola A, Dragic T, Arthos J, et al. (1996) CD4-dependent, antibody-sensitive interactions between HIV-1 and its co-receptor CCR-5. Nature 384: 184-187. doi: 10.1038/384184a0
|
| [106] |
Wyatt R, Sodroski J (1998) The HIV-1 envelope glycoproteins: fusogens, antigens, and immunogens. Science 280: 1884-1888. doi: 10.1126/science.280.5371.1884
|
| [107] |
Buzon V, Natrajan G, Schibli D, et al. (2010) Crystal structure of HIV-1 gp41 including both fusion peptide and membrane proximal external regions. PLoS Pathog 6: e1000880. doi: 10.1371/journal.ppat.1000880
|
| [108] |
Chan DC, Fass D, Berger JM, et al. (1997) Core structure of gp41 from the HIV envelope glycoprotein. Cell 89: 263-273. doi: 10.1016/S0092-8674(00)80205-6
|
| [109] |
Tan K, Liu J, Wang J, et al. (1997) Atomic structure of a thermostable subdomain of HIV-1 gp41. Proc Natl Acad Sci U S A 94: 12303-12308. doi: 10.1073/pnas.94.23.12303
|
| [110] |
Weissenhorn W, Dessen A, Harrison SC, et al. (1997) Atomic structure of the ectodomain from HIV-1 gp41. Nature 387: 426-430. doi: 10.1038/387426a0
|
| [111] |
Chen L, Kwon YD, Zhou T, et al. (2009) Structural basis of immune evasion at the site of CD4 attachment on HIV-1 gp120. Science 326: 1123-1127. doi: 10.1126/science.1175868
|
| [112] |
Huang CC, Lam SN, Acharya P, et al. (2007) Structures of the CCR5 N terminus and of a tyrosine-sulfated antibody with HIV-1 gp120 and CD4. Science 317: 1930-1934. doi: 10.1126/science.1145373
|
| [113] |
Huang CC, Tang M, Zhang MY, et al. (2005) Structure of a V3-containing HIV-1 gp120 core. Science 310: 1025-1028. doi: 10.1126/science.1118398
|
| [114] |
Huang J, Ofek G, Laub L, et al. (2012) Broad and potent neutralization of HIV-1 by a gp41-specific human antibody. Nature 491: 406-412. doi: 10.1038/nature11544
|
| [115] |
Kwong PD, Wyatt R, Robinson J, et al. (1998) Structure of an HIV gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody. Nature 393: 648-659. doi: 10.1038/31405
|
| [116] |
McLellan JS, Pancera M, Carrico C, et al. (2011) Structure of HIV-1 gp120 V1/V2 domain with broadly neutralizing antibody PG9. Nature 480: 336-343. doi: 10.1038/nature10696
|
| [117] |
Pejchal R, Doores KJ, Walker LM, et al. (2011) A potent and broad neutralizing antibody recognizes and penetrates the HIV glycan shield. Science 334: 1097-1103. doi: 10.1126/science.1213256
|
| [118] |
Ward AB, Wilson IA (2015) Insights into the trimeric HIV-1 envelope glycoprotein structure. Trends Biochem Sci 40: 101-107. doi: 10.1016/j.tibs.2014.12.006
|
| [119] |
Zhou T, Georgiev I, Wu X, et al. (2010) Structural basis for broad and potent neutralization of HIV-1 by antibody VRC01. Science 329: 811-817. doi: 10.1126/science.1192819
|
| [120] |
Zhou T, Xu L, Dey B, et al. (2007) Structural definition of a conserved neutralization epitope on HIV-1 gp120. Nature 445: 732-737. doi: 10.1038/nature05580
|
| [121] |
Dutta M, Liu J, Roux KH, et al. (2014) Visualization of retroviral envelope spikes in complex with the V3 loop antibody 447-52D on intact viruses by cryo-electron tomography. J Virol 88: 12265-12275. doi: 10.1128/JVI.01596-14
|
| [122] |
Harris AK, Bartesaghi A, Milne JL, et al. (2013) HIV-1 envelope glycoprotein trimers display open quaternary conformation when bound to the gp41 membrane-proximal external-region-directed broadly neutralizing antibody Z13e1. J Virol 87: 7191-7196. doi: 10.1128/JVI.03284-12
|
| [123] |
Hu G, Liu J, Taylor KA, et al. (2011) Structural comparison of HIV-1 envelope spikes with and without the V1/V2 loop. J Virol 85: 2741-2750. doi: 10.1128/JVI.01612-10
|
| [124] |
Zhu P, Winkler H, Chertova E, et al. (2008) Cryoelectron tomography of HIV-1 envelope spikes: further evidence for tripod-like legs. PLoS Pathog 4: e1000203. doi: 10.1371/journal.ppat.1000203
|
| [125] |
Sanders RW, Vesanen M, Schuelke N, et al. (2002) Stabilization of the soluble, cleaved, trimeric form of the envelope glycoprotein complex of human immunodeficiency virus type 1. J Virol 76: 8875-8889. doi: 10.1128/JVI.76.17.8875-8889.2002
|
| [126] |
Earl LA, Lifson JD, Subramaniam S (2013) Catching HIV 'in the act' with 3D electron microscopy. Trends Microbiol 21: 397-404. doi: 10.1016/j.tim.2013.06.004
|
| [127] |
Lucic V, Rigort A, Baumeister W (2013) Cryo-electron tomography: the challenge of doing structural biology in situ. J Cell Biol 202: 407-419. doi: 10.1083/jcb.201304193
|
| [128] |
Jun S, Ke D, Debiec K, et al. (2011) Direct visualization of HIV-1 with correlative live-cell microscopy and cryo-electron tomography. Structure 19: 1573-1581. doi: 10.1016/j.str.2011.09.006
|
| [129] | Jun S, Zhao G, Ning J, et al. (2013) Correlative microscopy for 3D structural analysis of dynamic interactions. J Vis Exp. |
| [130] |
Majorovits E, Nejmeddine M, Tanaka Y, et al. (2008) Human T-lymphotropic virus-1 visualized at the virological synapse by electron tomography. PLoS One 3: e2251. doi: 10.1371/journal.pone.0002251
|
| [131] |
Martin N, Welsch S, Jolly C, et al. (2010) Virological synapse-mediated spread of human immunodeficiency virus type 1 between T cells is sensitive to entry inhibition. J Virol 84: 3516-3527. doi: 10.1128/JVI.02651-09
|
| [132] |
Do T, Murphy G, Earl LA, et al. (2014) Three-dimensional imaging of HIV-1 virological synapses reveals membrane architectures involved in virus transmission. J Virol 88: 10327-10339. doi: 10.1128/JVI.00788-14
|
| [133] | Drobne D (2013) 3D imaging of cells and tissues by focused ion beam/scanning electron microscopy (FIB/SEM). Methods Mol Biol 950: 275-292. |
| [134] |
Risco C, Castro IFd, Sanz-Sánchez L, et al. (2014) Three-Dimensional Imaging of Viral Infections. Annual Review of Virology 1: 453-473. doi: 10.1146/annurev-virology-031413-085351
|
| [135] |
Kukulski W, Schorb M, Welsch S, et al. (2011) Correlated fluorescence and 3D electron microscopy with high sensitivity and spatial precision. J Cell Biol 192: 111-119. doi: 10.1083/jcb.201009037
|
| [136] |
Sartori A, Gatz R, Beck F, et al. (2007) Correlative microscopy: bridging the gap between fluorescence light microscopy and cryo-electron tomography. J Struct Biol 160: 135-145. doi: 10.1016/j.jsb.2007.07.011
|
| [137] |
Zhang P (2013) Correlative cryo-electron tomography and optical microscopy of cells. Curr Opin Struct Biol 23: 763-770. doi: 10.1016/j.sbi.2013.07.017
|
| [138] | Woodward CL, Cheng SN, Jensen GJ (2014) Electron cryo-tomography studies of maturing HIV-1 particles reveal the assembly pathway of the viral core. J Virol. |
| [139] |
Carlson LA, de Marco A, Oberwinkler H, et al. (2010) Cryo electron tomography of native HIV-1 budding sites. PLoS Pathog 6: e1001173. doi: 10.1371/journal.ppat.1001173
|
| [140] |
Dobro MJ, Samson RY, Yu Z, et al. (2013) Electron cryotomography of ESCRT assemblies and dividing Sulfolobus cells suggests that spiraling filaments are involved in membrane scission. Mol Biol Cell 24: 2319-2327. doi: 10.1091/mbc.E12-11-0785
|
| [141] |
Effantin G, Dordor A, Sandrin V, et al. (2013) ESCRT-III CHMP2A and CHMP3 form variable helical polymers in vitro and act synergistically during HIV-1 budding. Cell Microbiol 15: 213-226. doi: 10.1111/cmi.12041
|
| [142] |
Shen QT, Schuh AL, Zheng Y, et al. (2014) Structural analysis and modeling reveals new mechanisms governing ESCRT-III spiral filament assembly. J Cell Biol 206: 763-777. doi: 10.1083/jcb.201403108
|
| [143] |
Van Engelenburg SB, Shtengel G, Sengupta P, et al. (2014) Distribution of ESCRT machinery at HIV assembly sites reveals virus scaffolding of ESCRT subunits. Science 343: 653-656. doi: 10.1126/science.1247786
|
| [144] |
de Marco A, Muller B, Glass B, et al. (2010) Structural analysis of HIV-1 maturation using cryo-electron tomography. PLoS Pathog 6: e1001215. doi: 10.1371/journal.ppat.1001215
|
| [145] |
de Marco A, Heuser AM, Glass B, et al. (2012) Role of the SP2 domain and its proteolytic cleavage in HIV-1 structural maturation and infectivity. J Virol 86: 13708-13716. doi: 10.1128/JVI.01704-12
|
| [146] |
Konnyu B, Sadiq SK, Turanyi T, et al. (2013) Gag-Pol processing during HIV-1 virion maturation: a systems biology approach. PLoS Comput Biol 9: e1003103. doi: 10.1371/journal.pcbi.1003103
|
| [147] |
Keller PW, Adamson CS, Heymann JB, et al. (2011) HIV-1 maturation inhibitor bevirimat stabilizes the immature Gag lattice. J Virol 85: 1420-1428. doi: 10.1128/JVI.01926-10
|
| [148] |
Benjamin J, Ganser-Pornillos BK, Tivol WF, et al. (2005) Three-dimensional structure of HIV-1 virus-like particles by electron cryotomography. J Mol Biol 346: 577-588. doi: 10.1016/j.jmb.2004.11.064
|
| [149] |
Levandovsky A, Zandi R (2009) Nonequilibirum assembly, retroviruses, and conical structures. Phys Rev Lett 102: 198102. doi: 10.1103/PhysRevLett.102.198102
|
| [150] |
Yu Z, Dobro MJ, Woodward CL, et al. (2013) Unclosed HIV-1 capsids suggest a curled sheet model of assembly. J Mol Biol 425: 112-123. doi: 10.1016/j.jmb.2012.10.006
|
| [151] |
Sougrat R, Bartesaghi A, Lifson JD, et al. (2007) Electron tomography of the contact between T cells and SIV/HIV-1: implications for viral entry. PLoS Pathog 3: e63. doi: 10.1371/journal.ppat.0030063
|
| [152] |
Cardone G, Brecher M, Fontana J, et al. (2012) Visualization of the two-step fusion process of the retrovirus avian sarcoma/leukosis virus by cryo-electron tomography. J Virol 86: 12129-12137. doi: 10.1128/JVI.01880-12
|
| [153] |
Felts RL, Narayan K, Estes JD, et al. (2010) 3D visualization of HIV transfer at the virological synapse between dendritic cells and T cells. Proc Natl Acad Sci U S A 107: 13336-13341. doi: 10.1073/pnas.1003040107
|
| [154] |
Sherer NM, Lehmann MJ, Jimenez-Soto LF, et al. (2007) Retroviruses can establish filopodial bridges for efficient cell-to-cell transmission. Nat Cell Biol 9: 310-315. doi: 10.1038/ncb1544
|
| [155] | Pique C, Jones KS (2012) Pathways of cell-cell transmission of HTLV-1. Front Microbiol 3: 378. |
| [156] |
Hill CP, Worthylake D, Bancroft DP, et al. (1996) Crystal structures of the trimeric human immunodeficiency virus type 1 matrix protein: implications for membrane association and assembly. Proc Natl Acad Sci U S A 93: 3099-3104. doi: 10.1073/pnas.93.7.3099
|
| [157] |
Rao Z, Belyaev AS, Fry E, et al. (1995) Crystal structure of SIV matrix antigen and implications for virus assembly. Nature 378: 743-747. doi: 10.1038/378743a0
|
| 1. | Siming Liu, Peng Hou, Yingkun Gao, Yong Tan, Innovation and green total factor productivity in China: a linear and nonlinear investigation, 2020, 0944-1344, 10.1007/s11356-020-11436-1 | |
| 2. | Amr Hosny, Nonresident holdings of domestic debt in Nigeria: Internal or external driven?, 2020, 32, 1017-6772, 472, 10.1111/1467-8268.12453 | |
| 3. | Zhenghui Li, Hao Dong, Zimei Huang, Pierre Failler, Impact of Foreign Direct Investment on Environmental Performance, 2019, 11, 2071-1050, 3538, 10.3390/su11133538 | |
| 4. | Durmuş Yıldırım, Ali Kemal Çelik, Testing the pecking order theory of capital structure: Evidence from Turkey using panel quantile regression approach, 2020, 22148450, 10.1016/j.bir.2020.11.002 | |
| 5. | Siming Liu, Xiaoyan Shen, Tianpei Jiang, Pierre Failler, Impacts of the financialization of manufacturing enterprises on total factor productivity: empirical examination from China's listed companies, 2021, 3, 2643-1092, 59, 10.3934/GF.2021005 | |
| 6. | Tinghui Li, Junhao Zhong, Hai Zhang, Pierre Failler, Chinese financial cycle spillovers to developed countries, 2019, 1, 2643-1092, 364, 10.3934/GF.2019.4.364 | |
| 7. | Zhenghui Li, Yan Wang, Yong Tan, Zimei Huang, Does Corporate Financialization Affect Corporate Environmental Responsibility? An Empirical Study of China, 2020, 12, 2071-1050, 3696, 10.3390/su12093696 | |
| 8. | Financial Innovation and Financial Inclusion Nexus in South Asian Countries: Evidence from Symmetric and Asymmetric Panel Investigation, 2019, 7, 2227-7072, 61, 10.3390/ijfs7040061 | |
| 9. | Tough Chinoda, The nexus between financial inclusion, trade and economic growth in Africa?, 2020, 12, 1918-6444, 266, 10.1080/19186444.2020.1742065 | |
| 10. | Zejun Li, Zimei Huang, Tinghui Li, Jie Yan, Does Economic Growth Driving Force Convert? Evidence from China, 2020, 2020, 1563-5147, 1, 10.1155/2020/2768025 | |
| 11. | Sa Xu, Cunyi Yang, Zhehao Huang, Pierre Failler, Interaction between Digital Economy and Environmental Pollution: New Evidence from a Spatial Perspective, 2022, 19, 1660-4601, 5074, 10.3390/ijerph19095074 | |
| 12. | Immaculate Simiso Nxumalo, Patricia Lindelwa Makoni, Determinants of Foreign Capital Inflows in Emerging Markets: The Role of Institutional Quality, 2021, 7, 2518-8488, 683, 10.26710/jafee.v7i3.1881 | |
| 13. | Yeasmin Islam, Piana Monsur Mindia, Nusrat Farzana, Md Qamruzzaman, Nexus between environmental sustainability, good governance, financial inclusion, and tourism development in Bangladesh: Evidence from symmetric and asymmetric investigation, 2023, 10, 2296-665X, 10.3389/fenvs.2022.1056268 | |
| 14. | Kashif Islam, Ahmad Raza Bilal, Syed Anees Haider Zaidi, Symmetric and asymmetric nexus between economic freedom and stock market development in Pakistan, 2022, 55, 1573-9414, 2391, 10.1007/s10644-022-09385-5 | |
| 15. | Yaya Su, Zhenghui Li, Cunyi Yang, Spatial Interaction Spillover Effects between Digital Financial Technology and Urban Ecological Efficiency in China: An Empirical Study Based on Spatial Simultaneous Equations, 2021, 18, 1660-4601, 8535, 10.3390/ijerph18168535 | |
| 16. | Zheng Shi, Md. Qamruzzaman, Re-Visiting the Role of Education on Poverty Through the Channel of Financial Inclusion: Evidence From Lower-Income and Lower-Middle-Income Countries, 2022, 10, 2296-665X, 10.3389/fenvs.2022.873652 | |
| 17. | Yixing Yang, Md. Qamruzzaman, Mohd Ziaur Rehman, Salma Karim, Do Tourism and Institutional Quality Asymmetrically Effects on FDI Sustainability in BIMSTEC Countries: An Application of ARDL, CS-ARDL, NARDL, and Asymmetric Causality Test, 2021, 13, 2071-1050, 9989, 10.3390/su13179989 | |
| 18. | Kexian Zhang, Xiaoying Liu, Min Hong, László Vasa, Discretionary Effort on Green Technology Innovation: How Chinese Enterprises Act when Facing Financing Constraints, 2021, 16, 1932-6203, e0261589, 10.1371/journal.pone.0261589 | |
| 19. | Long JinRu, Md Qamruzzaman, Nexus Between Environmental Innovation, Energy Efficiency, and Environmental Sustainability in G7: What is the Role of Institutional Quality?, 2022, 10, 2296-665X, 10.3389/fenvs.2022.860244 | |
| 20. | Jianxin Zhuo, Md Qamruzzaman, Do financial development, FDI, and globalization intensify environmental degradation through the channel of energy consumption: evidence from belt and road countries, 2022, 29, 0944-1344, 2753, 10.1007/s11356-021-15796-0 | |
| 21. | Long JinRu, Md. Qamruzzaman, Wu Hangyu, Rajnish Kler, Do environmental quality, financial inclusion, and good governance ensure the FDI sustainably in Belt and Road countries? Evidence from an application of CS-ARDL and NARDL, 2022, 10, 2296-665X, 10.3389/fenvs.2022.936216 | |
| 22. | Mustafa Hassan Mohammad Adam, Nexus among foreign direct investment, financial development, and sustainable economic growth: Empirical aspects from Sudan, 2022, 6, 2573-0134, 640, 10.3934/QFE.2022028 | |
| 23. | Dito Rinaldo, Vina Anggilia Puspita, 2021, 978-1-80117-595-1, 169, 10.1108/S1571-03862021000029A025 | |
| 24. | Osman ALTAY, SOVEREIGN-BANK NEXUS OF TURKEY: AN ANALYSIS OF SOVEREIGN RISK AND BANKING INDUSTRY PERFORMANCE INDICATORS BASED ON CAMELS RATING SYSTEM, 2021, 22, 1302-6739, 217, 10.31671/doujournal.973012 | |
| 25. | Piotr Dąbrowski, Stock Indices Breakdown during the Pandemic as the Most Dynamic Bear Market in History: Consequences for Individual Investors, 2021, 10, 2227-9091, 1, 10.3390/risks10010001 | |
| 26. | Guang Liu, Yunying Huang, Zhehao Huang, Determinants and Mechanisms of Digital Financial Inclusion Development: Based on Urban-Rural Differences, 2021, 11, 2073-4395, 1833, 10.3390/agronomy11091833 | |
| 27. | Lijin Zhao, Md. Qamruzzaman, Do Urbanization, Remittances, and Globalization Matter for Energy Consumption in Belt and Road Countries: Evidence From Renewable and Non-Renewable Energy Consumption, 2022, 10, 2296-665X, 10.3389/fenvs.2022.930728 | |
| 28. | Urgaia R. Worku, Factor productivity nexus economic growth in Sub-Saharan Africa: Symmetric and asymmetric panel approaches, 2024, 5, 2772655X, 100169, 10.1016/j.wds.2024.100169 | |
| 29. | Sofía Orazi, Lisana B. Martinez, Hernán P. Vigier, Determinants and evolution of financial inclusion in Latin America: A demand side analysis, 2023, 7, 2573-0134, 187, 10.3934/QFE.2023010 | |
| 30. | Raed Walid Al-Smadi, Arkan Walid Al-Smadi, The relationship between foreign direct investment and financial inclusion in MENA countries: Evidence from the General Method of Moments, 2024, 21, 18104967, 144, 10.21511/imfi.21(2).2024.11 | |
| 31. | Md. Qamruzzaman, Simon Grima, Does financial innovation foster financial inclusion in Arab world? examining the nexus between financial innovation, FDI, remittances, trade openness, and gross capital formation, 2023, 18, 1932-6203, e0287475, 10.1371/journal.pone.0287475 | |
| 32. | Mohammed Musah, Stephen Taiwo Onifade, Isaac Ankrah, Bright Akwasi Gyamfi, George Kofi Amoako, Achieving net-zero emission target in Africa: Are sustainable energy innovations and financialization crucial for environmental sustainability of sub-Saharan African state?, 2024, 364, 03062619, 123120, 10.1016/j.apenergy.2024.123120 | |
| 33. | Asuman Balat, Levent Çıtak, FİNANSAL GELİŞMENİN DOĞRUDAN YABANCI YATIRIM VE YABANCI PORTFÖY YATIRIMI ÜZERİNDEKİ ETKİSİ: TÜRKİYE ÜZERİNE BİR UYGULAMA, 2024, 2687-5306, 10.56574/nohusosbil.1560316 | |
| 34. | Omar Ikbal Tawfik, Mohammed Ali Ahmed, Hamada Elsaid Elmaasrawy, The Mediating Role of Mobile Banking-Based Financial Inclusion Disclosure on the Relationship Between Foreign Investment and Bank Performance, 2024, 12, 2227-7072, 128, 10.3390/ijfs12040128 |
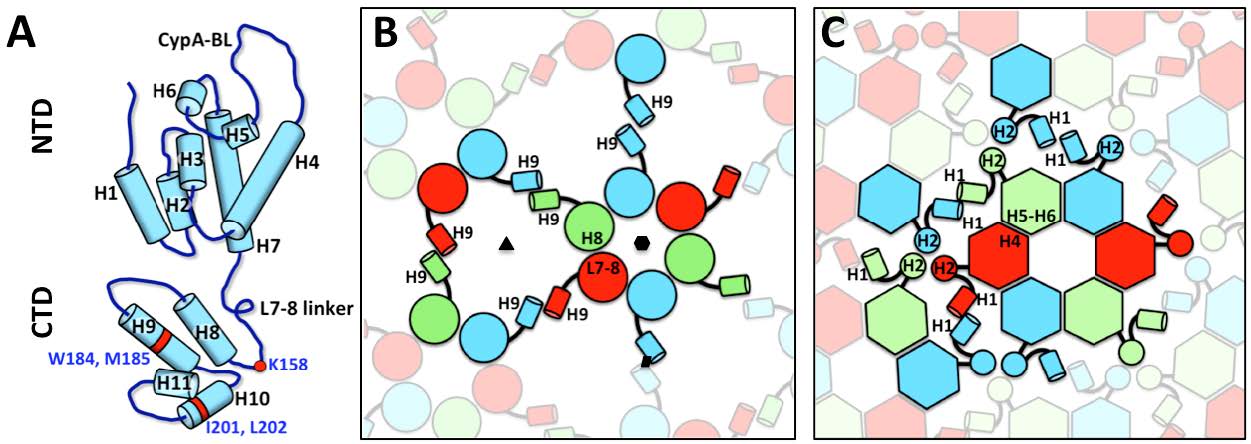
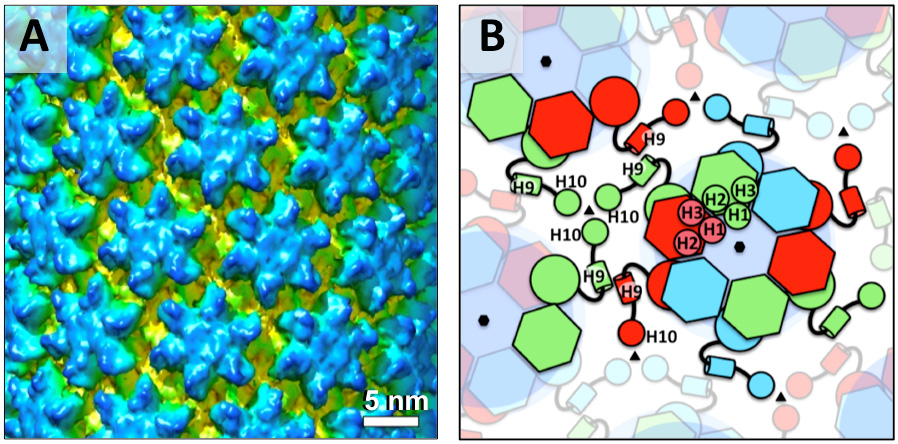
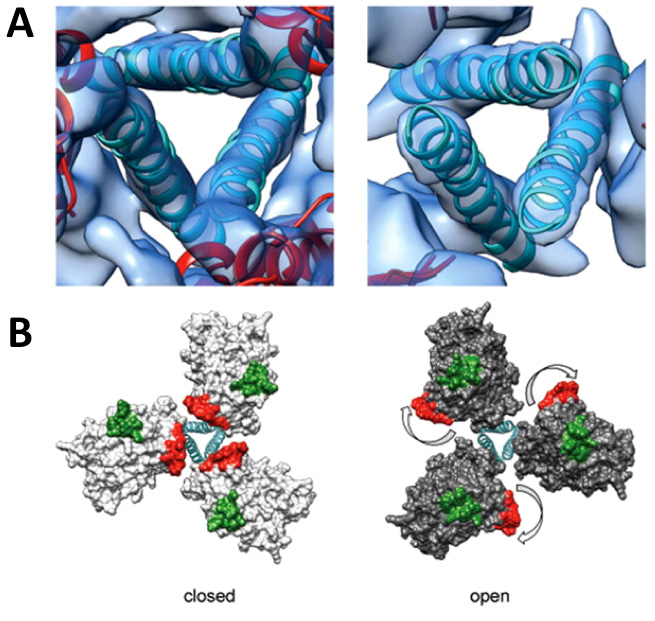
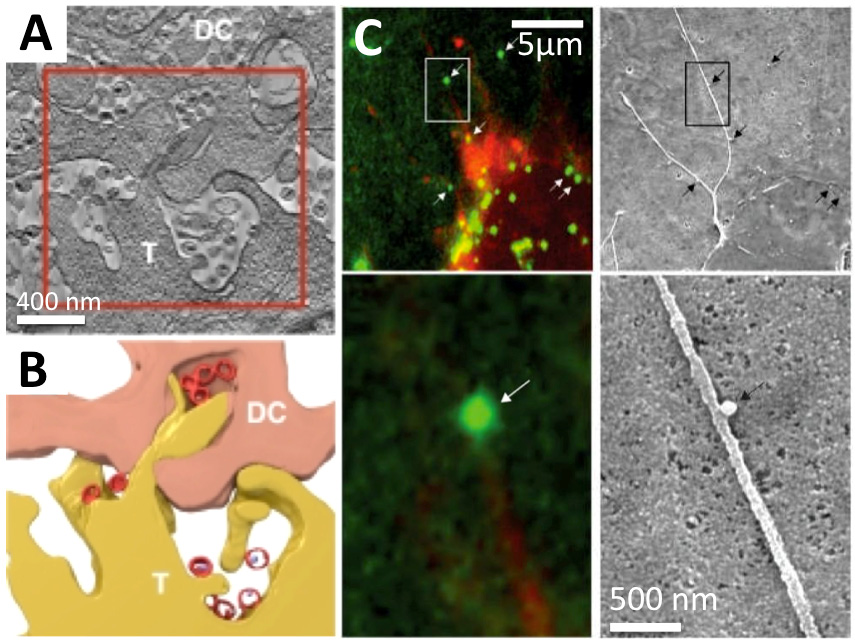
Figures(7) / Tables(1)

Wei Zhang, Sheng Cao, Jessica L. Martin, Joachim D. Mueller, Louis M. Mansky. Morphology and ultrastructure of retrovirus particles[J]. AIMS Biophysics, 2015, 2(3): 343-369. doi: 10.3934/biophy.2015.3.343

| Factor | Conversion | Yield of DPE | Selectivity towards DPE |
| Temperature °C | P = 0.0502 | P = 0.3526 | P = 0.3785 |
| Type of Catalyst | P = 0.0012 | P = 0.0312 | P = 0.0249 |
| P < 0.05 indicates significance | |||
DownLoad: CSV| Responses | ||||||
| Factor | Conversion of Methanol | Conversion of n-propanol | Yield of DPE | Yield of MPE | Selectivity towards DPE | Selectivity toward MPE |
| Temperature, °C | P =<0.0001 | P =<0.0001 | P =<0.0001 | P = 0.595 | P =<0.0001 | P =<0.0001 |
| Type of Catalyst | P = 0.0003 | P =<0.0001 | P =<0.0001 | P = 0.9 | P = 0.0181 | P = 0.0181 |
| Catalyst Concentration | P =<0.0001 | P =<0.0001 | P = 0.0010 | P = 0.0295 | P = 0.0295 | P = 0.0295 |
| Catalyst Concentration *Type of Catalyst | P = 0.0181 | P = 0.0089 | P = 0.0176 | P = 0.8815 | P = 0.2027 | P = 0.2027 |
| *P: Probability value for significance. P < 0.05 indicates significance. | ||||||
DownLoad: CSV| Factor | Conversion | Yield of DPE | Selectivity towards DPE |
| Temperature °C | P = 0.0502 | P = 0.3526 | P = 0.3785 |
| Type of Catalyst | P = 0.0012 | P = 0.0312 | P = 0.0249 |
| P < 0.05 indicates significance | |||
| Responses | ||||||
| Factor | Conversion of Methanol | Conversion of n-propanol | Yield of DPE | Yield of MPE | Selectivity towards DPE | Selectivity toward MPE |
| Temperature, °C | P =<0.0001 | P =<0.0001 | P =<0.0001 | P = 0.595 | P =<0.0001 | P =<0.0001 |
| Type of Catalyst | P = 0.0003 | P =<0.0001 | P =<0.0001 | P = 0.9 | P = 0.0181 | P = 0.0181 |
| Catalyst Concentration | P =<0.0001 | P =<0.0001 | P = 0.0010 | P = 0.0295 | P = 0.0295 | P = 0.0295 |
| Catalyst Concentration *Type of Catalyst | P = 0.0181 | P = 0.0089 | P = 0.0176 | P = 0.8815 | P = 0.2027 | P = 0.2027 |
| *P: Probability value for significance. P < 0.05 indicates significance. | ||||||