Citation: Arindam Mitra, Suman Mukhopadhyay. Biofilm mediated decontamination of pollutants from the environment[J]. AIMS Bioengineering, 2016, 3(1): 44-59. doi: 10.3934/bioeng.2016.1.44
| [1] | Yan Pan, Fuyong Jiao . Challenges in early diagnosis and treatment of Kawasaki disease. AIMS Allergy and Immunology, 2024, 8(4): 213-215. doi: 10.3934/Allergy.2024012 |
| [2] | Florentina-Silvia Delli, Elena Sotiriou, Efstratios Vakirlis, Demetrios Ioannides . Bullous pemphigoid autoantibodies. AIMS Allergy and Immunology, 2021, 5(4): 259-263. doi: 10.3934/Allergy.2021019 |
| [3] | Mansur Aliyu, Ali Akbar Saboor-Yaraghi, Shima Nejati, Behrouz Robat-Jazi . Urinary VPAC1: A potential biomarker in prostate cancer. AIMS Allergy and Immunology, 2022, 6(2): 42-63. doi: 10.3934/Allergy.2022006 |
| [4] | Hazhar M. Balaky, Parween Abdulsamad Ismail . Serum resistin, visfatin, IL-17 and IL-23 as novel diagnostic biomarkers for thyroid carcinoma. AIMS Allergy and Immunology, 2025, 9(3): 166-179. doi: 10.3934/Allergy.2025013 |
| [5] | Darrell O. Ricke . Antibodies and infected monocytes and macrophages in COVID-19 patients. AIMS Allergy and Immunology, 2022, 6(2): 64-70. doi: 10.3934/Allergy.2022007 |
| [6] | Raghda S. El Mashaari, Ameera S. Al Ali, Zaidoon M. Abdel Hadi, Mahmoud Talaat Sabri, Ahmed Nageeb Mahmoud . COVID-19 vaccine-induced new-onset pustular psoriasis mimicking skin infection of the hand and foot: A report of two cases. AIMS Allergy and Immunology, 2025, 9(2): 136-143. doi: 10.3934/Allergy.2025010 |
| [7] | Haiyan Guo, Mingsheng Lei, Jinan Ma, Hongchun Du, Youming Zhang . Allergy and ferroptosis. AIMS Allergy and Immunology, 2025, 9(1): 8-26. doi: 10.3934/Allergy.2025002 |
| [8] | Tuba Karakurt, Nurhan Kasap, Kübra Aslan, Hayrunnisa Bekis Bozkurt, Fatma Bal Cetinkaya, Gizem Uslu, Zafer Bicakci, Filiz Özen, Özlem Cavkaytar, Ahmet Eken, Mustafa Arga . Suspected ALPS with clinical and laboratory findings: Three patients—three different diagnoses. AIMS Allergy and Immunology, 2023, 7(4): 304-312. doi: 10.3934/Allergy.2023020 |
| [9] | Hui-Min Zhang, Qi-Ling Yin, You-Qiong Liu, Ya-Le Zhang, Wei-Hua Zhang . Recurrent Kawasaki disease in children: Four case reports. AIMS Allergy and Immunology, 2025, 9(2): 123-135. doi: 10.3934/Allergy.2025009 |
| [10] | Muhammad Sadeqi Nezhad, Farhad Seif, Ilad Alavi Darazam, Azam Samei, Monireh Kamali, Hossein Aazami, Monireh Mohsenzadegan, Yaghoub Mollaei-Kandelousi, Pegah Babaheidarian, Majid Khoshmirsafa, Mohsen Fateh . An overview of the prominence of current diagnostic methods for diagnosis of COVID-19. AIMS Allergy and Immunology, 2020, 4(3): 60-74. doi: 10.3934/Allergy.2020006 |
| Aβ: amyloid-β protein | AD: Alzheimer's disease |
| aPLs: antiphospholipid autoantibodies | APP: amyloid-β precursor protein |
| BBB: blood-brain barrier | CDR: complementarity determining region |
| CNS: central nervous system | CSF: cerebrospinal fluid |
| Ig: immunoglobulin | FcRn: neonatal fragment crystallizable region (of Ig) receptor |
| MAPτ: microtubule-associated protein τ | MCI: mild cognitive impairment |
| PD: Parkinson's disease | R-RAA: redox-reactive autoantibody |
The diagnosis "Alzheimer's disease" originally described a hereditary form of early onset dementia [1] with a characteristic, postmortem, histopathological correlate in the central nervous tissue of neurofibrillary tangles, amyloid plaques and neuron loss. Nowadays, "Alzheimer's disease" includes conditions hitherto known as "senile dementia" because of the similar histopathology showing shrinkage of cerebral cortex and hippocampus, enlarged ventricles and amyloid deposits [2]. Diagnostic criteria of disease are regularly reviewed to provide clinical guidelines [3]. Alzheimer's disease (AD) is a chronic fatal neurodegenerative disorder whose prevalence is on the rise. The most recent estimate is that there are 47 million AD patients in the world [4]. The impact on society is significant, not only financially in terms of health care and other provisions, but also with regard to the sustainability of intergenerational relationships when cognitive loss impoverishes traditions and community cohesions. Current treatment targets levels of neurotransmitters and slows progression of disease [5]. The diagnosis of AD is verified postmortem on identification of neurofibrillary tangles formed by τ protein and senile plaques formed by aggregated amyloid-β protein (Aβ). There is a need for an accurate and easy method to establish the diagnosis of AD early.
Biomarkers (ie measures which reflect activity of disease) have been identified for AD, such as reduced levels of cellular protein Aβ, increased levels of normally intracellular total τ protein and its phosphorylated form p-τ, which are measurable in cerebrospinal fluid (CSF) [6]. A blood test, however, has not yet been developed that shows comparable sensitivity and specificity for the probable diagnosis of AD.
Autoantibodies are normally present at low titers. Their pathogenic involvement is strongly suggested when titers increase during chronic inflammatory disease with organ damage. D'Andrea (2003) showed greater reactivity of neuron death in brain parenchyma with neuronal-specific anti-immunoglobulin (Ig) antibodies and a breach in blood brain barrier (BBB) that had occurred in AD patients [7]. Through leakage of plasma proteins or upregulation in inherent production of complement components, the classical complement pathway may be engaged in the brains of AD patients, sustaining inflammation and activation of microglia. This may lead to enhanced antigen presentation and processing of potential autoantigens [8]. Contrasting with other biomarkers made evident by AD damage, the change of autoantibody amounts may precede the damage to patients' cognition. Therefore, this review pursues the hypothesis that circulating autoantibodies in AD relate to disease and may have the potential to be a biomarker for early stage AD diagnosis.
AD entails nervous lesions in brain. This means that the disease-related molecules are located in the central nervous system (CNS) and/or CSF, escaping easy detection. Autoantibodies can build the bridge from internal brain molecules to blood by crossing the BBB. Normally, the BBB secures an environment specific to the CNS, which is suspended in CSF, an ultrafiltrate of plasma produced by the choroid plexus located in some of the brain ventricles. Although autoantibodies can enter CSF temporarily, they are normally shuttled out via the neonatal Fc Receptor (FcRn) [9]. Once its capacity is exceeded, however, presence of significant autoantibody levels against τ protein and heavy neurofilament may be detected in CSF from AD patients [10]. Ageing can induce breakdown of BBB with a predilection for the hippocampus, leading to early stage AD or mild cognitive impairment (MCI) [11]. Most cerebropathy-related autoantibodies are transported by blood circulation other than the immigration of effector B cells [12]. So, the concentration of autoantibody in blood is relative to the AD-associated damage in brain. This discovery can be seen as the theoretical basis of blood-based autoantibody biomarker detection.
The complementarity determining regions (CDR) of IgG may bind to AD pathology-related molecules, neurotransmitters and receptors, glial markers, phospholipids and so on. Those autoantibodies recognizing Aβ protein and τ protein seem to be more specific and reliable than other autoantibodies because these molecules can link to AD directly and apparently change pre-symptomatically. The roles that autoantibodies play in AD are different, even totally opposite from one to another. Some of them cause damage of the normal tissue or amplify the damages in AD patients' brain similar to autoimmune disease, based on the colocalization of antibody reactivity with neuronal death [7]; others recognize abnormal host molecules and may try to eliminate them as a kind of defense from a severe AD, as seen from Aβ clearance by FcRn mediated transcytosis, which increases with age, across the BBB [13]. This means that the increase and decrease of the autoantibody can be quite different with regard to the functional impacts.
Aβs originate from a transmembrane protein amyloid-β precursor protein (APP). APPs are ubiquitous in health with unknown function. Aβs also have a wide range of isoforms, but Aβ40 and Aβ42 (where 40 and 42 indicate the numbers of amino acid residues) are the most prevalent in the human body. The abnormally high production and accumulation of Aβs, especially Aβ42, in amyloid neurotic plaques and cerebral vessels in brain are most likely to be the pathogenic pathway accompanying AD. Aβ accumulation is capable of inducing inflammation of glial cells [14] and can block synaptic transmission inside CNS [15]. Aβ itself is not a useful biomarker in the diagnosis of AD. The main problem for that is the multitudinous states of Aβ, such as monomer Aβ, oligomer and polymer Aβ accumulation [16], protein complex bound Aβ [17], and of immune complex formation with autoantibody [18]. These states of Aβ not only have different molecular weights and different biochemical features, but also mutually transform continuously in the early stage of AD, so no single current quantitative biological technique can gauge the exact gross amount of Aβ.
Comparing with Aβ, Anti-Aβ autoantibodies only have two states: bound and unbound. As mentioned, anti-Aβ autoantibodies can be detected in blood samples. The quantitative change of anti-Aβ autoantibodies has been debated for a long time. Some studies suggested that the anti-Aβ autoantibodies in AD patients are higher than the healthy controls [19,20], while other scientists hold the statement that anti-Aβ autoantibodies decrease in AD sufferers [21]. The sample sizes in these studies were rather small. Moreover, Li et al. (2007) proved that employing low pH agent to dissociate the autoantibody overestimates levels [22]. Gustaw-Rothenberg et al. (2010) have investigated a large, age matched controlled, sample set randomly selected from over two million inhabitants from the Lublin Region in Poland [23]. In the first year of diagnosis of AD, the number of autoantibodies, especially the non-dissociated type, increased dramatically. But after the first year of AD progression, this number dropped gradually below normal. This result showed that anti-Aβ autoantibodies have the potential to be utilized as a biomarker for the early stage AD or MCI. It can be explained that the autoantibodies can only bind to the oligo-Aβ other than mono-Aβ or Aβ accumulation deposit. Aβ engages in a variety of multiple complexes in the CSF of AD patients' brain, resulting in the significant decrease of the concentration of mono-Aβs [17,24]. Gruden et al. (2007) suggested the level of anti-Aβ autoantibody is down-regulated by a fall of oligo-Aβ forms which have accumulated into polymer-Aβ accumulation or are bound with antibody [25]. Because of the ability of FcRn to transport Aβ immune complexes from brain to blood, anti Aβ antibodies may function as a peripheral sink [26] and detection of anti-Aβ autoantibody will not mirror these complex dynamics of the disease process.
T protein is the cleavage product of microtubule-associated protein τ (MAPτ), a part of the microtubular cytoskeleton in CNS. Topical in the studies of neurological diseases such as AD and Parkinson's disease (PD), it is suspected as another important AD pathology-associated molecule with a different pathomechanism from Aβ. The τ protein has many potential phosphorylation sites, if these sites are hyperphosphorylated during inflammation in CSF and brain in AD patients, this abnormal structure results in the self-assembly of tangles of paired helical filaments and straight filaments which lead to neuronal damage [27].
The studies of anti-τ antibodies are not as numerous as those of anti-Aβ. Anti-τ antibodies were shown to increase in AD patients [28]. Interestingly, in contrast to the AD patients, the anti-τ antibodies in other kinds of dementia patients decrease substantially [10]. Hromadkova et al. (2015) showed that there are anti-τ reactive antibodies present in pooled fractions of IgG from thousands of healthy donors [29]. A subsequent study screened various types of serum autoantibodies by τ deposits in vitro, and found that the selected anti-τ antibodies recognized τ tangles and threads and inhibited τ aggregation [30]. With such evidence, anti-τ antibodies may play a defensive role in AD.
S100b is a glial marker. S100bs are secreted by a particular astrocyte ensheathing vessels and by NG2 (Neuron-glial antigen 2)-expressing cells, which may be oligodendrocyte precursors [31]. As an astrocyte-derived cytokine, the alteration of S100b can be used as a biomarker for several kinds of dementias involving astrocyte activation [32]. As astrocyte foot processes form part of the physical BBB, any disturbance of astrocyte viability compromises the BBB.
The role S100b plays in AD remains a puzzle. In a mouse model, overexpression of S100b (which may be induced by interleukin-1 [33]) correlated with synthesis of neuritic plaques [34]. Another report pointed out S100b is essential in keeping the permeability of BBB in wildtype mice and that S100b knock out mice showed dysfunction of endothelial cells [35]. On the other hand, the amount change of anti-S100b autoantibodies in AD patients can agree with each other. Studies show the anti-S100b antibodies increase in MCI phase and decrease in severe AD phase, as described for anti-Aβ autoantibodies [36]. Gruden et al. (2007) examined four kinds of autoantibodies which target at Aβ, S100b and two kinds of neurotransmitters in the same sample [25]. The increase and decrease of anti-Aβ and anti-S100b autoantibodies were synchronistic. This result implied that at least S100b and Aβ have some notable or potential relation to each other.
Among the antiphospholipid autoantibodies (aPLs), there is a recently characterised family called redox-reactive autoantibodies (R-RAAs) which holds promise to be a biomarker for AD diagnosis [37,38]. R-RAAs recognize specifically self-antigens subsequent to redox reactions, in other words, they are sensitive to reaction endproducts after exposure to transitional metals (manganese or iron) in CSF, blood or other internal environment [39]. Therefore, it is easy to separate R-RAAs from other natural and hidden autoantibodies by immunoassay [40]. In fact, the key symptom of AD is the neurodegenerative disorder. Neurons, particularly sensitive to oxidative stress, are vulnerable to undergo apoptosis or necrosis [41]. Neuritic plaques induce an abundant metal redox agent environment in CSF [42]. Moreover, the tyrosine residues in the CDRs of unmasking redox autoantibodies are nitrosylated at the same time of neurotic plaque formation [38]. In immunoassay examination of CSF, R-RAAs are significantly decreased in autopsy-confirmed AD patients compared to healthy controls [43]. R-RAAs appear to be a reverse biomarker that decreases substantially with the increase in severity of AD, so, conversely, they may be very useful to diagnose AD in the early stage. Recently, a study was set up to compare the amount of R-RAAs among age matched groups of healthy controls, patients with MCI and AD [44]. Interestingly, like other autoantibodies mentioned before, the result showed that the amount of R-RAAs rose in MCI patients and dropped in the severe AD patients relative to the controls. This rise may relate to the hemin (iron containing porphyrin) recruitment in the early stage of AD. Then, with the development of disease, these autoantibodies will lose the hemin environment and their titre decreases. Like anti-τ proteins, R-RAA are reported to decrease in the other dementia patients [45].
A good biomarker should have 3 key features: high specificity (to exclude those not diseased), high sensitivity (to diagnose the diseased) and be convenient to analyze. Although most autoantibody biomarkers can be detected in blood-based samples, which is cheaper and easier than obtaining a CSF sample, it is not advisable to depend on only one specific kind of autoantibody as a biomarker for AD diagnosis. There are two reasons for that: for the autoantibodies whose targets are not pathology-specific molecules, their targets are usually not unique in AD but also can be found in other cerebropathy or neuropathy; even if the autoantibodies against Aβ or autoantibodies against τ, their concentrations change over a large range over time intervals in one patient or vary considerably between patients in a group. To solve this problem, the diagnosis can set multiple biomarkers as combination detection, or additional clinical diagnostics to improve the sensitivity and specificity. This, in principle, is being pursued when combining the measurement of CSF biomarkers with diagnostic imaging [46,47].
From the elaborations made so far, anti-Aβ, anti-S100b and anti-phospholipid autoantibodies are of interest for early stage AD diagnosis, as they have an interesting rise in the stage of MIC (Table 1).
| Name of auto-antibody | Function to AD | Quantitative change in stage of MCI | Quantitative change in advanced stage of AD | Exclusion from other causes of dementia |
| Anti-Aβ | Neutral | Increase | Decrease | Yes |
| Anti-τ | Negative | Unknown | Increase | Yes |
| Anti-S100b | Unknown | Increase | Decrease | Unknown |
| Anti-phospholipid | Unknown | Increase | Decrease | No |
 DownLoad: CSV
DownLoad: CSVPresently there are two schools of thought, i. to pursue the identification of AD specific autoantibodies using protein microarray or phage display technology with a view to derive pathomechanistic understanding and candidates for therapeutic targeting [48,49], ii. to focus on the discovery of disease phase characteristic antibodies in AD, in clear distinction of healthy controls, using synthetic peptoid libraries, where the natural ligand remains unknown [50].
While the former aids in the appreciation of dynamics in AD related autoantibody levels (MCI vs severe stage), the latter approach seems of particular promise in order to further diagnosis in the early symptomatic, MCI, phase of AD, thereby allowing timeliness of adjustments and interventions [51], irrespective of exact pathomechanistic understanding.
The first author is recipient of the President's Postgraduate Scholarship Scheme at the University of Leicester.
The authors have no conflict of interest to declare.
| [1] | Alexander M, Loehr RC (1992) Bioremediation review. Science 258: 874. |
| [2] | Prasad MN, Prasad R (2012) Nature's cure for cleanup of contaminated environment—a review of bioremediation strategies. Rev Environ Health 27: 181–189. |
| [3] |
Vidali M (2001) Bioremediation. an overview. Pure and Applied Chemistry 73: 1163–1172. doi: 10.1351/pac200173071163

|
| [4] |
Gaur N, Flora G, Yadav M, et al. (2014) A review with recent advancements on bioremediation-based abolition of heavy metals. Environ Sci Process Impacts 16: 180–193. doi: 10.1039/C3EM00491K
|
| [5] |
Atlas RM, Hazen TC (2011) Oil biodegradation and bioremediation: a tale of the two worst spills in U.S. history. Environ Sci Technol 45: 6709–6715. doi: 10.1021/es2013227
|
| [6] |
Bragg JR, Prince RC, Harner EJ, et al. (1994) Effectiveness of bioremediation for the Exxon Valdez oil spill. Nature 368: 413–418. doi: 10.1038/368413a0
|
| [7] |
Day SM (1993) US environmental regulations and policies--their impact on the commercial development of bioremediation. Trends Biotechnol 11: 324–328. doi: 10.1016/0167-7799(93)90154-2
|
| [8] |
Caplan JA (1993) The worldwide bioremediation industry: prospects for profit. Trends Biotechnol 11: 320–323. doi: 10.1016/0167-7799(93)90153-Z
|
| [9] |
Mishra A, Malik A (2014) Novel fungal consortium for bioremediation of metals and dyes from mixed waste stream. Bioresour Technol 171: 217–226. doi: 10.1016/j.biortech.2014.08.047
|
| [10] |
Cerniglia CE (1997) Fungal metabolism of polycyclic aromatic hydrocarbons: past, present and future applications in bioremediation. J Ind Microbiol Biotechnol 19: 324–333. doi: 10.1038/sj.jim.2900459
|
| [11] | Balaji V, Arulazhagan P, Ebenezer P (2014) Enzymatic bioremediation of polyaromatic hydrocarbons by fungal consortia enriched from petroleum contaminated soil and oil seeds. J Environ Biol 35: 521–529. |
| [12] |
Bouwer EJ, Zehnder AJ (1993) Bioremediation of organic compounds--putting microbial metabolism to work. Trends Biotechnol 11: 360–367. doi: 10.1016/0167-7799(93)90159-7
|
| [13] |
Bruins MR, Kapil S, Oehme FW (2000) Microbial resistance to metals in the environment. Ecotoxicol Environ Saf 45: 198–207. doi: 10.1006/eesa.1999.1860
|
| [14] | Prince RC (2000) Bioremediation. Kirk-Othmer Encyclopedia of Chemical Technology: John Wiley & Sons, Inc. |
| [15] | Das S, Dash HR (2014) 1 - Microbial Bioremediation: A Potential Tool for Restoration of Contaminated Areas. In: Das S, editor. Microbial Biodegradation and Bioremediation. Oxford: Elsevier. pp. 1–21. |
| [16] | Joutey NT, Sayel H, Bahafid W, et al. (2015) Mechanisms of hexavalent chromium resistance and removal by microorganisms. Rev Environ Contam Toxicol 233: 45–69. |
| [17] |
Kumar R, Singh S, Singh OV (2007) Bioremediation of radionuclides: emerging technologies. OMICS 11: 295–304. doi: 10.1089/omi.2007.0013
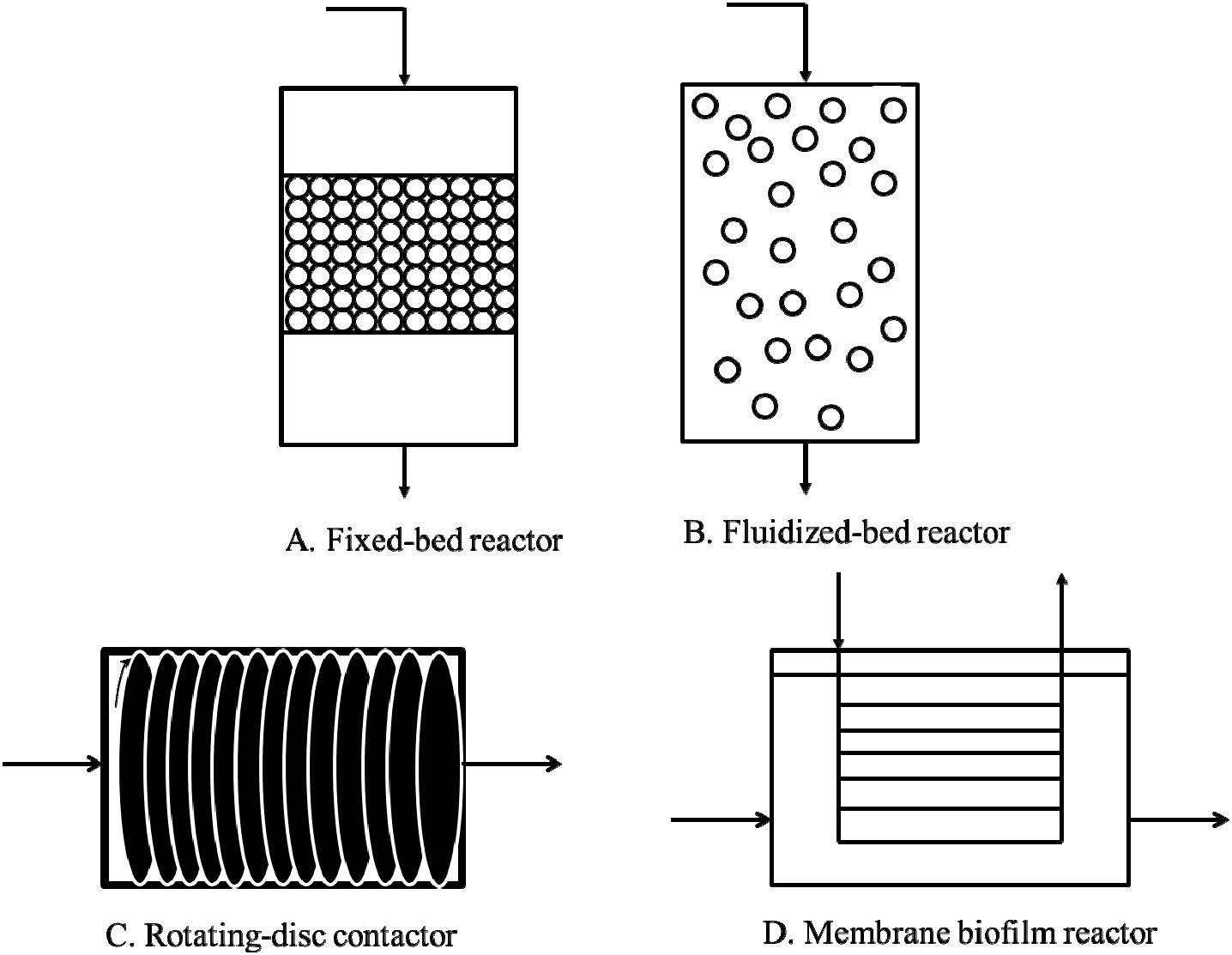
|
| [18] |
Wall JD, Krumholz LR (2006) Uranium reduction. Annu Rev Microbiol 60: 149–166. doi: 10.1146/annurev.micro.59.030804.121357
|
| [19] |
Beyenal H, Sani RK, Peyton BM, et al. (2004) Uranium immobilization by sulfate-reducing biofilms. Environ Sci Technol 38: 2067–2074. doi: 10.1021/es0348703
|
| [20] | Vogt C, Richnow HH (2014) Bioremediation via in situ microbial degradation of organic pollutants. Adv Biochem Eng Biotechnol 142: 123–146. |
| [21] |
Jorgensen KS (2007) In situ bioremediation. Adv Appl Microbiol 61: 285–305. doi: 10.1016/S0065-2164(06)61008-3
|
| [22] |
Singh JS, Abhilash PC, Singh HB, et al. (2011) Genetically engineered bacteria: an emerging tool for environmental remediation and future research perspectives. Gene 480: 1–9. doi: 10.1016/j.gene.2011.03.001
|
| [23] | Hedlund BP, Staley JT (2001) Vibrio cyclotrophicus sp. nov., a polycyclic aromatic hydrocarbon (PAH)-degrading marine bacterium. Int J Syst Evol Microbiol 51: 61–66. |
| [24] |
Nakajima-Kambe T, Ichihashi F, Matsuzoe R, et al. (2009) Degradation of aliphatic–aromatic copolyesters by bacteria that can degrade aliphatic polyesters. Polym Degrad Stab 94: 1901–1905. doi: 10.1016/j.polymdegradstab.2009.08.006
|
| [25] |
Costerton JW, Cheng KJ, Geesey GG, et al. (1987) Bacterial biofilms in nature and disease. Annu Rev Microbiol 41: 435–464. doi: 10.1146/annurev.mi.41.100187.002251
|
| [26] |
Gieg LM, Fowler SJ, Berdugo-Clavijo C (2014) Syntrophic biodegradation of hydrocarbon contaminants. Curr Opin Biotechnol 27: 21–29. doi: 10.1016/j.copbio.2013.09.002
|
| [27] |
Horemans B, Breugelmans P, Hofkens J, et al. (2013) Environmental dissolved organic matter governs biofilm formation and subsequent linuron degradation activity of a linuron-degrading bacterial consortium. Appl Environ Microbiol 79: 4534–4542. doi: 10.1128/AEM.03730-12
|
| [28] |
Pratt LA, Kolter R (1999) Genetic analyses of bacterial biofilm formation. Curr Opin Microbiol 2: 598–603. doi: 10.1016/S1369-5274(99)00028-4
|
| [29] |
Lacal J, Reyes-Darias JA, García-Fontana C, et al. (2013) Tactic responses to pollutants and their potential to increase biodegradation efficiency. J Appl Microbiol 114: 923–933. doi: 10.1111/jam.12076
|
| [30] | Flemming HC, Wingender J (2010) The biofilm matrix. Nat Rev Microbiol 8: 623–633. |
| [31] |
More TT, Yadav JSS, Yan S, et al. (2014) Extracellular polymeric substances of bacteria and their potential environmental applications. J Environ Manage 144: 1–25. doi: 10.1016/j.jenvman.2014.05.010
|
| [32] | Flemming HC, Wingender J (2001) Relevance of microbial extracellular polymeric substances (EPSs)--Part II: Technical aspects. Water Sci Technol 43: 9–16. |
| [33] |
Branda SS, Vik S, Friedman L, et al. (2005) Biofilms: the matrix revisited. Trends Microbiol 13: 20–26. doi: 10.1016/j.tim.2004.11.006
|
| [34] |
Jung JH, Choi NY, Lee SY (2013) Biofilm formation and exopolysaccharide (EPS) production by Cronobacter sakazakii depending on environmental conditions. Food Microbiol 34: 70–80. doi: 10.1016/j.fm.2012.11.008
|
| [35] | Kreft JU, Wimpenny JW (2001) Effect of EPS on biofilm structure and function as revealed by an individual-based model of biofilm growth. Water Sci Technol 43: 135–141. |
| [36] |
Miqueleto AP, Dolosic CC, Pozzi E, et al. (2010) Influence of carbon sources and C/N ratio on EPS production in anaerobic sequencing batch biofilm reactors for wastewater treatment. Bioresour Technol 101: 1324–1330. doi: 10.1016/j.biortech.2009.09.026
|
| [37] |
Reysenbach AL, Cady SL (2001) Microbiology of ancient and modern hydrothermal systems. Trends Microbiol 9: 79–86. doi: 10.1016/S0966-842X(00)01921-1
|
| [38] |
Edwards KJ, Bond PL, Gihring TM, et al. (2000) An archaeal iron-oxidizing extreme acidophile important in acid mine drainage. Science 287: 1796–1799. doi: 10.1126/science.287.5459.1796
|
| [39] |
Matz C, Kjelleberg S (2005) Off the hook--how bacteria survive protozoan grazing. Trends Microbiol 13: 302–307. doi: 10.1016/j.tim.2005.05.009
|
| [40] |
Davey ME, O'Toole G A (2000) Microbial biofilms: from ecology to molecular genetics. Microbiol Mol Biol Rev 64: 847–867. doi: 10.1128/MMBR.64.4.847-867.2000
|
| [41] |
Mah TF, O'Toole GA (2001) Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol 9: 34–39. doi: 10.1016/S0966-842X(00)01913-2
|
| [42] |
Sutherland IW (2001) The biofilm matrix--an immobilized but dynamic microbial environment. Trends Microbiol 9: 222–227. doi: 10.1016/S0966-842X(01)02012-1
|
| [43] |
Field JA, Stams AJ, Kato M, et al. (1995) Enhanced biodegradation of aromatic pollutants in cocultures of anaerobic and aerobic bacterial consortia. Antonie Van Leeuwenhoek 67: 47–77. doi: 10.1007/BF00872195
|
| [44] |
De Philippis R, Colica G, Micheletti E (2011) Exopolysaccharide-producing cyanobacteria in heavy metal removal from water: molecular basis and practical applicability of the biosorption process. Appl Microbiol Biotechnol 92: 697–708. doi: 10.1007/s00253-011-3601-z
|
| [45] |
De Philippis R, Paperi R, Sili C (2007) Heavy metal sorption by released polysaccharides and whole cultures of two exopolysaccharide-producing cyanobacteria. Biodegradation 18: 181–187. doi: 10.1007/s10532-006-9053-y
|
| [46] |
Micheletti E, Colica G, Viti C, et al. (2008) Selectivity in the heavy metal removal by exopolysaccharide-producing cyanobacteria. J Appl Microbiol 105: 88–94. doi: 10.1111/j.1365-2672.2008.03728.x
|
| [47] |
Iwabuchi N, Sunairi M, Urai M, et al. (2002) Extracellular polysaccharides of Rhodococcus rhodochrous S-2 stimulate the degradation of aromatic components in crude oil by indigenous marine bacteria. Appl Environ Microbiol 68: 2337–2343. doi: 10.1128/AEM.68.5.2337-2343.2002
|
| [48] |
Li W-W, Yu H-Q (2014) Insight into the roles of microbial extracellular polymer substances in metal biosorption. Bioresource Technology 160: 15–23. doi: 10.1016/j.biortech.2013.11.074
|
| [49] |
Pal A, Paul AK (2008) Microbial extracellular polymeric substances: central elements in heavy metal bioremediation. Indian J Microbiol 48: 49–64. doi: 10.1007/s12088-008-0006-5
|
| [50] | Ferris FG, Schultze S, Witten TC, et al. (1989) Metal Interactions with Microbial Biofilms in Acidic and Neutral pH Environments. Appl Environ Microbiol 55: 1249–1257. |
| [51] |
Zhang HL, Fang W, Wang YP, et al. (2013) Phosphorus removal in an enhanced biological phosphorus removal process: roles of extracellular polymeric substances. Environ Sci Technol 47: 11482–11489. doi: 10.1021/es403227p
|
| [52] |
Yuan Z, Pratt S, Batstone DJ (2012) Phosphorus recovery from wastewater through microbial processes. Curr Opin Biotechnol 23: 878–883. doi: 10.1016/j.copbio.2012.08.001
|
| [53] |
Burmolle M, Webb JS, Rao D, et al. (2006) Enhanced biofilm formation and increased resistance to antimicrobial agents and bacterial invasion are caused by synergistic interactions in multispecies biofilms. Appl Environ Microbiol 72: 3916–3923. doi: 10.1128/AEM.03022-05
|
| [54] |
Logsdon GS, Kohne R, Abel S, et al. (2002) Slow sand filtration for small water systems. J Environ Eng Sci 1: 339–348. doi: 10.1139/s02-025
|
| [55] | Kartal B, Kuenen JG, van Loosdrecht MC (2010) Engineering. Sewage treatment with anammox. Science 328: 702–703. |
| [56] |
Bengtsson MM, Ovreas L (2010) Planctomycetes dominate biofilms on surfaces of the kelp Laminaria hyperborea. BMC Microbiol 10: 261. doi: 10.1186/1471-2180-10-261
|
| [57] | Fuchs S, Haritopoulou T, Schäfer M, et al. (1997) Heavy metals in freshwater ecosystems introduced by urban rainwater runoff—Monitoring of suspended solids, river sediments and biofilms. Water Sci Technol 36: 277–282. |
| [58] | Peacock AD, Chang YJ, Istok JD, et al. (2004) Utilization of microbial biofilms as monitors of bioremediation. Microb Ecol 47: 284–292. |
| [59] |
Brummer IH, Fehr W, Wagner-Dobler I (2000) Biofilm community structure in polluted rivers: abundance of dominant phylogenetic groups over a complete annual cycle. Appl Environ Microbiol 66: 3078–3082. doi: 10.1128/AEM.66.7.3078-3082.2000
|
| [60] |
Arini A, Feurtet–Mazel A, Maury-Brachet R, et al. (2012) Recovery potential of periphytic biofilms translocated in artificial streams after industrial contamination (Cd and Zn). Ecotoxicology 21: 1403–1414. doi: 10.1007/s10646-012-0894-3
|
| [61] |
SzabÓ KÉ, Makk J, Kiss KT, et al. (2008) Sequential colonization by river periphyton analysed by microscopy and molecular fingerprinting. Freshwater Biology 53: 1359–1371. doi: 10.1111/j.1365-2427.2008.01967.x
|
| [62] |
Dorigo U, Bérard A, Humbert JF (2002) Comparison of Eukaryotic Phytobenthic Community Composition in a Polluted River by Partial 18S rRNA Gene Cloning and Sequencing. Microbial Ecology 44: 372–380. doi: 10.1007/s00248-002-2024-x
|
| [63] |
Kostanjsek R, Lapanje A, Drobne D, et al. (2005) Bacterial community structure analyses to assess pollution of water and sediments in the Lake Shkodra/Skadar, Balkan Peninsula. Environ Sci Pollut Res Int 12: 361–368. doi: 10.1065/espr2005.07.271
|
| [64] |
Bricheux G, Le Moal G, Hennequin C, et al. (2013) Characterization and evolution of natural aquatic biofilm communities exposed in vitro to herbicides. Ecotoxicol Environ Saf 88: 126–134. doi: 10.1016/j.ecoenv.2012.11.003
|
| [65] |
Ancion PY, Lear G, Dopheide A, et al. (2013) Metal concentrations in stream biofilm and sediments and their potential to explain biofilm microbial community structure. Environ Pollut 173: 117–124. doi: 10.1016/j.envpol.2012.10.012
|
| [66] |
Navarro E, Guasch H, Sabater S (2002) Use of microbenthic algal communities in ecotoxicological tests for the assessment of water quality: the Ter river case study. J Appl Phycol 14: 41–48. doi: 10.1023/A:1015242301451
|
| [67] |
Dorigo U, Bourrain X, Bérard A, et al. (2004) Seasonal changes in the sensitivity of river microalgae to atrazine and isoproturon along a contamination gradient. Sci Total Environ 318: 101–114. doi: 10.1016/S0048-9697(03)00398-X
|
| [68] |
Dewez D, Didur O, Vincent-Héroux J, et al. (2008) Validation of photosynthetic-fluorescence parameters as biomarkers for isoproturon toxic effect on alga Scenedesmus obliquus. Environ Pollution 151: 93–100. doi: 10.1016/j.envpol.2007.03.002
|
| [69] |
Schmitt-Jansen M, Altenburger R (2008) Community-level microalgal toxicity assessment by multiwavelength-excitation PAM fluorometry. Aquat Toxicol 86: 49–58. doi: 10.1016/j.aquatox.2007.10.001
|
| [70] |
Sayler GS, Layton A, Lajoie C, et al. (1995) Molecular site assessment and process monitoring in bioremediation and natural attenuation. off. Appl Biochem Biotechnol 54: 277–290. doi: 10.1007/BF02787926
|
| [71] |
Jorgensen KS, Salminen JM, Bjorklof K (2010) Monitored natural attenuation. Methods Mol Biol 599: 217–233. doi: 10.1007/978-1-60761-439-5_14
|
| [72] |
Rittmann BE (2004) Definition, objectives, and evaluation of natural attenuation. Biodegradation 15: 349–357. doi: 10.1023/B:BIOD.0000044587.05189.99
|
| [73] |
Tyagi M, da Fonseca MM, de Carvalho CC (2011) Bioaugmentation and biostimulation strategies to improve the effectiveness of bioremediation processes. Biodegradation 22: 231–241. doi: 10.1007/s10532-010-9394-4
|
| [74] |
Jansson JK, Bjorklof K, Elvang AM, et al. (2000) Biomarkers for monitoring efficacy of bioremediation by microbial inoculants. Environ Pollut 107: 217–223. doi: 10.1016/S0269-7491(99)00140-2
|
| [75] |
Gentry T, Rensing C, Pepper IAN (2004) New Approaches for Bioaugmentation as a Remediation Technology. Crit Rev Environ Sci Technol 34: 447–494. doi: 10.1080/10643380490452362
|
| [76] |
Morgan P, Watkinson RJ (1989) Hydrocarbon degradation in soils and methods for soil biotreatment. Crit Rev Biotechnol 8: 305–333. doi: 10.3109/07388558909148196
|
| [77] |
Grace Liu P-W, Chang TC, Whang L-M, et al. (2011) Bioremediation of petroleum hydrocarbon contaminated soil: Effects of strategies and microbial community shift. Int Biodeterior Biodegradation 65: 1119–1127. doi: 10.1016/j.ibiod.2011.09.002
|
| [78] |
Abeysinghe DH, De Silva DG, Stahl DA, et al. (2002) The effectiveness of bioaugmentation in nitrifying systems stressed by a washout condition and cold temperature. Water Environ Res 74: 187–199. doi: 10.2175/106143002X139901
|
| [79] |
Boon N, De Gelder L, Lievens H, et al. (2002) Bioaugmenting bioreactors for the continuous removal of 3-chloroaniline by a slow release approach. Environ Sci Technol 36: 4698–4704. doi: 10.1021/es020076q
|
| [80] |
Qureshi N, Annous BA, Ezeji TC, et al. (2005) Biofilm reactors for industrial bioconversion processes: employing potential of enhanced reaction rates. Microb Cell Fact 4: 24. doi: 10.1186/1475-2859-4-24
|
| [81] |
Bryers JD (1993) Bacterial biofilms. Curr Opin Biotechnol 4: 197–204. doi: 10.1016/0958-1669(93)90125-G
|
| [82] |
Rosche B, Li XZ, Hauer B, et al. (2009) Microbial biofilms: a concept for industrial catalysis? Trends Biotechnol 27: 636–643. doi: 10.1016/j.tibtech.2009.08.001
|
| [83] |
Singh R, Paul D, Jain RK (2006) Biofilms: implications in bioremediation. Trends Microbiol 14: 389–397. doi: 10.1016/j.tim.2006.07.001
|
| [84] |
Wagner-Dobler I (2003) Pilot plant for bioremediation of mercury-containing industrial wastewater. Appl Microbiol Biotechnol 62: 124–133. doi: 10.1007/s00253-003-1322-7
|
| [85] | Shieh W, Keenan J (1986) Fluidized bed biofilm reactor for wastewater treatment. Bioproducts: Springer Berlin Heidelberg. pp. 131–169. |
| [86] |
Denac M, Dunn IJ (1988) Packed- and fluidized-bed biofilm reactor performance for anaerobic wastewater treatment. Biotechnol Bioeng 32: 159–173. doi: 10.1002/bit.260320206
|
| [87] |
Kumar TA, Saravanan S (2009) Treatability studies of textile wastewater on an aerobic fluidized bed biofilm reactor (FABR): a case study. Water Sci Technol 59: 1817–1821. doi: 10.2166/wst.2009.207
|
| [88] |
Costley SC, Wallis FM (2001) Bioremediation of heavy metals in a synthetic wastewater using a rotating biological contactor. Water Res 35: 3715–3723. doi: 10.1016/S0043-1354(01)00072-0
|
| [89] |
Eker S, Kargi F (2008) Biological treatment of 2,4-dichlorophenol containing synthetic wastewater using a rotating brush biofilm reactor. Bioresour Technol 99: 2319–2325. doi: 10.1016/j.biortech.2007.05.016
|
| [90] |
Eker S, Kargi F (2010) COD, para-chlorophenol and toxicity removal from synthetic wastewater using rotating tubes biofilm reactor (RTBR). Bioresour Technol 101: 9020–9024. doi: 10.1016/j.biortech.2010.07.003
|
| [91] | Abraham TE, Senan RC, Shaffiqu TS, et al. (2003) Bioremediation of textile azo dyes by an aerobic bacterial consortium using a rotating biological contactor. Biotechnol Prog 19: 1372–1376. |
| [92] |
Jeswani H, Mukherji S (2012) Degradation of phenolics, nitrogen-heterocyclics and polynuclear aromatic hydrocarbons in a rotating biological contactor. Bioresour Technol 111: 12–20. doi: 10.1016/j.biortech.2012.01.157
|
| [93] |
Sarayu K, Sandhya S (2012) Rotating biological contactor reactor with biofilm promoting mats for treatment of benzene and xylene containing wastewater. Appl Biochem Biotechnol 168: 1928–1937. doi: 10.1007/s12010-012-9908-0
|
| [94] | Rittmann BE (2006) The membrane biofilm reactor: the natural partnership of membranes and biofilm. Water Sci Technol 53: 219–225. |
| [95] | Nerenberg R, Rittmann BE (2004) Hydrogen-based, hollow-fiber membrane biofilm reactor for reduction of perchlorate and other oxidized contaminants. Water Sci Technol 49: 223–230. |
| [96] |
Modin O, Fukushi K, Yamamoto K (2008) Simultaneous removal of nitrate and pesticides from groundwater using a methane-fed membrane biofilm reactor. Water Sci Technol 58: 1273–1279. doi: 10.2166/wst.2008.481
|
| [97] | Fathepure BZ, Vogel TM (1991) Complete degradation of polychlorinated hydrocarbons by a two-stage biofilm reactor. Appl Environ Microbiol 57: 3418–3422. |
| [98] |
Zhang C, Wang L, Yan N, et al. (2013) Air-lift internal loop biofilm reactor for realized simultaneous nitrification and denitrification. Bioprocess Biosyst Eng 36: 597–602. doi: 10.1007/s00449-012-0814-1
|
| [99] |
Zhao Y, Feng C, Wang Q, et al. (2011) Nitrate removal from groundwater by cooperating heterotrophic with autotrophic denitrification in a biofilm–electrode reactor. J Hazard Mater 192: 1033–1039. doi: 10.1016/j.jhazmat.2011.06.008
|
| [100] |
White C, Gadd GM (1998) Accumulation and effects of cadmium on sulphate-reducing bacterial biofilms. Microbiology 144: 1407–1415. doi: 10.1099/00221287-144-5-1407
|
| [101] |
White C, Gadd GM (2000) Copper accumulation by sulfate-reducing bacterial biofilms. FEMS Microbiology Letters 183: 313–318. doi: 10.1111/j.1574-6968.2000.tb08977.x
|
| [102] |
Smith WL, Gadd GM (2000) Reduction and precipitation of chromate by mixed culture sulphate-reducing bacterial biofilms. J Appl Microbiol 88: 983–991. doi: 10.1046/j.1365-2672.2000.01066.x
|
| [103] |
Hosseini Koupaie E, Alavi Moghaddam MR, Hashemi SH (2013) Evaluation of integrated anaerobic/aerobic fixed-bed sequencing batch biofilm reactor for decolorization and biodegradation of azo dye acid red 18: comparison of using two types of packing media. Bioresour Technol 127: 415–421. doi: 10.1016/j.biortech.2012.10.003
|
| [104] |
Lin YH, Hsien TY (2009) Kinetics of biodegradation of phenolic wastewater in a biofilm reactor. Water Sci Technol 59: 1703–1711. doi: 10.2166/wst.2009.203
|
| [105] |
Moreno-Andrade I, Buitron G, Vargas A (2009) Effect of starvation and shock loads on the biodegradation of 4-chlorophenol in a discontinuous moving bed biofilm reactor. Appl Biochem Biotechnol 158: 222–230. doi: 10.1007/s12010-008-8392-z
|
| [106] |
Coelhoso I, Boaventura R, Rodrigues A (1992) Biofilm reactors: an experimental and modeling study of wastewater denitrification in fluidized-bed reactors of activated carbon particles. Biotechnol Bioeng 40: 625–633. doi: 10.1002/bit.260400510
|
| [107] |
Masic A, Eberl HJ (2014) A modeling and simulation study of the role of suspended microbial populations in nitrification in a biofilm reactor. Bull Math Biol 76: 27–58. doi: 10.1007/s11538-013-9898-2
|
| [108] | Martin KJ, Picioreanu C, Nerenberg R (2015) Assessing microbial competition in a hydrogen-based membrane biofilm reactor (MBfR) using multidimensional modeling. Biotechnol Bioeng. |
| [109] |
Valls M, de Lorenzo Vc (2002) Exploiting the genetic and biochemical capacities of bacteria for the remediation of heavy metal pollution. FEMS Microbiol Rev 26: 327–338. doi: 10.1111/j.1574-6976.2002.tb00618.x
|
| [110] |
Diels L, De Smet M, Hooyberghs L, et al. (1999) Heavy metals bioremediation of soil. Molecular Biotechnology 12: 149–158. doi: 10.1385/MB:12:2:149
|
| [111] | Macaskie LE, Yong P, Doyle TC, et al. (1997) Bioremediation of uranium-bearing wastewater: biochemical and chemical factors influencing bioprocess application. Biotechnol Bioeng 53: 100–109. |
| [112] | Shukla SK, Mangwani N, Rao TS, et al. (2014) 8 - Biofilm-Mediated Bioremediation of Polycyclic Aromatic Hydrocarbons. In: Das S, editor. Microbial Biodegradation and Bioremediation. Oxford: Elsevier. pp. 203–232. |
| [113] | Chen M, Xu P, Zeng G, et al. (2015) Bioremediation of soils contaminated with polycyclic aromatic hydrocarbons, petroleum, pesticides, chlorophenols and heavy metals by composting: Applications, microbes and future research needs. Biotechnol Adv 33: 745–755. |
| [114] | Cerniglia C (1993) Biodegradation of polycyclic aromatic hydrocarbons. In: Rosenberg E, editor. Microorganisms to Combat Pollution: Springer Netherlands. pp. 227–244. |
| [115] |
Jones KC, de Voogt P (1999) Persistent organic pollutants (POPs): state of the science. Environ Pollut 100: 209–221. doi: 10.1016/S0269-7491(99)00098-6
|
| [116] |
Bonefeld-Jorgensen EC, Hjelmborg PS, Reinert TS, et al. (2006) Xenoestrogenic activity in blood of European and Inuit populations. Environ Health 5: 12. doi: 10.1186/1476-069X-5-12
|
| [117] | Johnsen AR, Karlson U (2004) Evaluation of bacterial strategies to promote the bioavailability of polycyclic aromatic hydrocarbons. Appl Microbiol Biotechnol 63: 452–459. |
| [118] |
Rodriguez S, Bishop P (2008) Enhancing the Biodegradation of Polycyclic Aromatic Hydrocarbons: Effects of Nonionic Surfactant Addition on Biofilm Function and Structure. J Environ Eng 134: 505–512. doi: 10.1061/(ASCE)0733-9372(2008)134:7(505)
|
| [119] |
Plosz BG, Vogelsang C, Macrae K, et al. (2010) The BIOZO process--a biofilm system combined with ozonation: occurrence of xenobiotic organic micro-pollutants in and removal of polycyclic aromatic hydrocarbons and nitrogen from landfill leachate. Water Sci Technol 61: 3188–3197. doi: 10.2166/wst.2010.920
|
| [120] | Song HG, Bartha R (1990) Effects of jet fuel spills on the microbial community of soil. Appl Environ Microbiol 56: 646–651. |
| [121] |
Ron EZ, Rosenberg E (2014) Enhanced bioremediation of oil spills in the sea. Curr Opin Biotechnol 27: 191–194. doi: 10.1016/j.copbio.2014.02.004
|
| [122] |
Harayama S, Kasai Y, Hara A (2004) Microbial communities in oil-contaminated seawater. Curr Opin Biotechnol 15: 205–214. doi: 10.1016/j.copbio.2004.04.002
|
| [123] | Dasgupta D, Ghosh R, Sengupta TK (2013) Biofilm-mediated enhanced crude oil degradation by newly isolated pseudomonas species. ISRN Biotechnol 2013: 250749. |
| [124] |
Mnif I, Mnif S, Sahnoun R, et al. (2015) Biodegradation of diesel oil by a novel microbial consortium: comparison between co-inoculation with biosurfactant-producing strain and exogenously added biosurfactants. Environ Sci Pollut Res Int 22: 14852–14861. doi: 10.1007/s11356-015-4488-5
|
| [125] |
Koren O, Knezevic V, Ron EZ, et al. (2003) Petroleum pollution bioremediation using water-insoluble uric acid as the nitrogen source. Appl Environ Microbiol 69: 6337–6339. doi: 10.1128/AEM.69.10.6337-6339.2003
|
| [126] | Muyzer G, Stams AJ (2008) The ecology and biotechnology of sulphate-reducing bacteria. Nat Rev Microbiol 6: 441–454. |
| [127] | Erable B, Duţeanu NM, Ghangrekar MM, et al. (2009) Application of electro-active biofilms. Biofouling 26: 57–71. |
| [128] |
Li Z, Zhang X, Lei L (2008) Electricity production during the treatment of real electroplating wastewater containing Cr6+ using microbial fuel cell. Process Biochemistry 43: 1352–1358. doi: 10.1016/j.procbio.2008.08.005
|
| [129] |
Cong Y, Xu Q, Feng H, et al. (2013) Efficient electrochemically active biofilm denitrification and bacteria consortium analysis. Bioresour Technol 132: 24–27. doi: 10.1016/j.biortech.2013.01.004
|
| [130] |
Heitzer A, Sayler GS (1993) Monitoring the efficacy of bioremediation. Trends Biotechnol 11: 334–343. doi: 10.1016/0167-7799(93)90156-4
|
| [131] |
Perumbakkam S, Hess TF, Crawford RL (2006) A bioremediation approach using natural transformation in pure-culture and mixed-population biofilms. Biodegradation 17: 545–557. doi: 10.1007/s10532-005-9025-7
|
| [132] |
Urgun-Demirtas M, Stark B, Pagilla K (2006) Use of Genetically Engineered Microorganisms (GEMs) for the Bioremediation of Contaminants. Crit Rev Biotechnol 26: 145–164. doi: 10.1080/07388550600842794
|
| [133] | Cases I, de Lorenzo V (2005) Genetically modified organisms for the environment: stories of success and failure and what we have learned from them. Int Microbiol 8: 213–222. |
| [134] | Absalon C, Ymele-Leki P, Watnick PI (2012) The bacterial biofilm matrix as a platform for protein delivery. MBio 3: e00127–00112. |
| [135] |
Safa M, Alemzadeh I, Vossoughi M (2014) Biodegradability of oily wastewater using rotating biological contactor combined with an external membrane. J Environ Health Sci Eng 12: 117. doi: 10.1186/s40201-014-0117-3
|
| [136] |
Kaindl N (2010) Upgrading of an activated sludge wastewater treatment plant by adding a moving bed biofilm reactor as pre-treatment and ozonation followed by biofiltration for enhanced COD reduction: design and operation experience. Water Sci Technol 62: 2710–2719. doi: 10.2166/wst.2010.938
|
| 1. | Miyo K. Chatanaka, Dorsa Sohaei, Eleftherios P. Diamandis, Ioannis Prassas, Beyond the amyloid hypothesis: how current research implicates autoimmunity in Alzheimer’s disease pathogenesis, 2023, 1040-8363, 1, 10.1080/10408363.2023.2187342 | |
| 2. | Dimitrina Miteva, Georgi V. Vasilev, Tsvetelina Velikova, Role of Specific Autoantibodies in Neurodegenerative Diseases: Pathogenic Antibodies or Promising Biomarkers for Diagnosis, 2023, 12, 2073-4468, 81, 10.3390/antib12040081 |
Figures(1)

Arindam Mitra, Suman Mukhopadhyay. Biofilm mediated decontamination of pollutants from the environment[J]. AIMS Bioengineering, 2016, 3(1): 44-59. doi: 10.3934/bioeng.2016.1.44

| Name of auto-antibody | Function to AD | Quantitative change in stage of MCI | Quantitative change in advanced stage of AD | Exclusion from other causes of dementia |
| Anti-Aβ | Neutral | Increase | Decrease | Yes |
| Anti-τ | Negative | Unknown | Increase | Yes |
| Anti-S100b | Unknown | Increase | Decrease | Unknown |
| Anti-phospholipid | Unknown | Increase | Decrease | No |
DownLoad: CSV| Name of auto-antibody | Function to AD | Quantitative change in stage of MCI | Quantitative change in advanced stage of AD | Exclusion from other causes of dementia |
| Anti-Aβ | Neutral | Increase | Decrease | Yes |
| Anti-τ | Negative | Unknown | Increase | Yes |
| Anti-S100b | Unknown | Increase | Decrease | Unknown |
| Anti-phospholipid | Unknown | Increase | Decrease | No |





