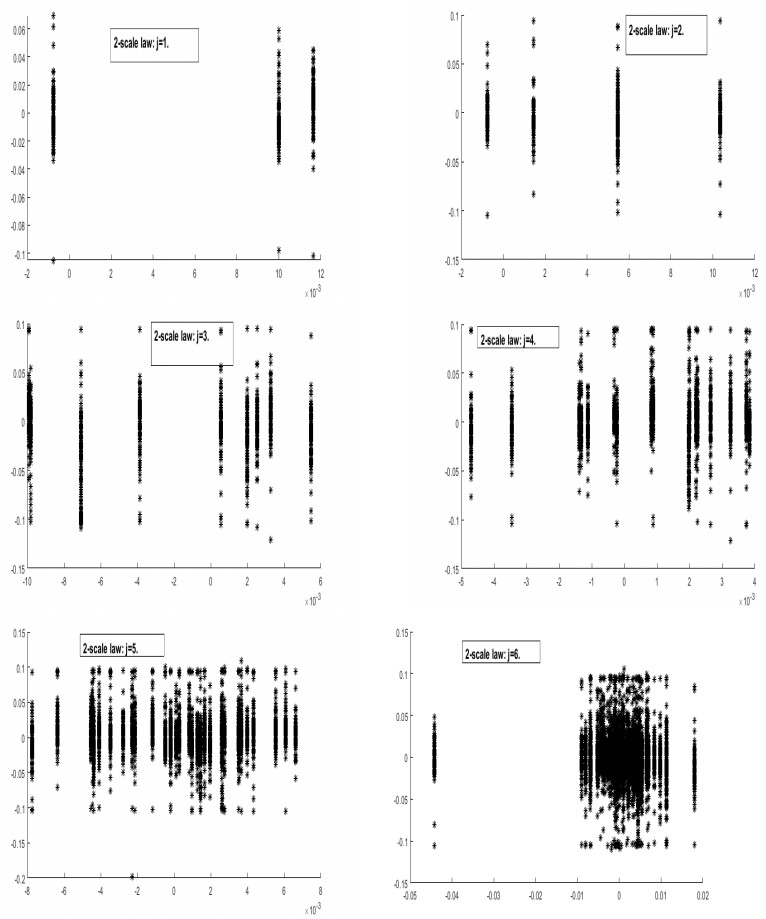
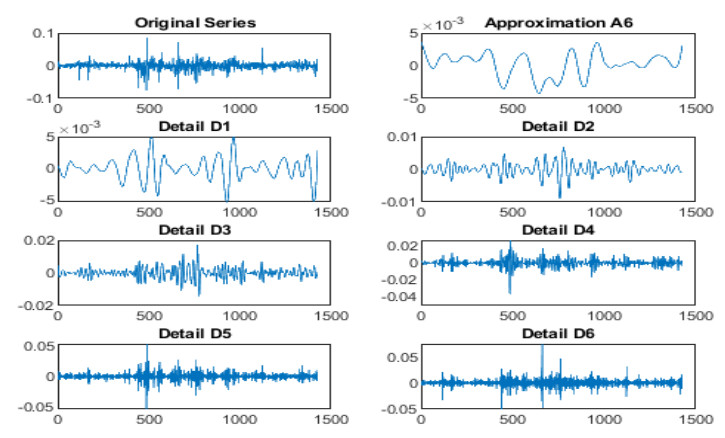
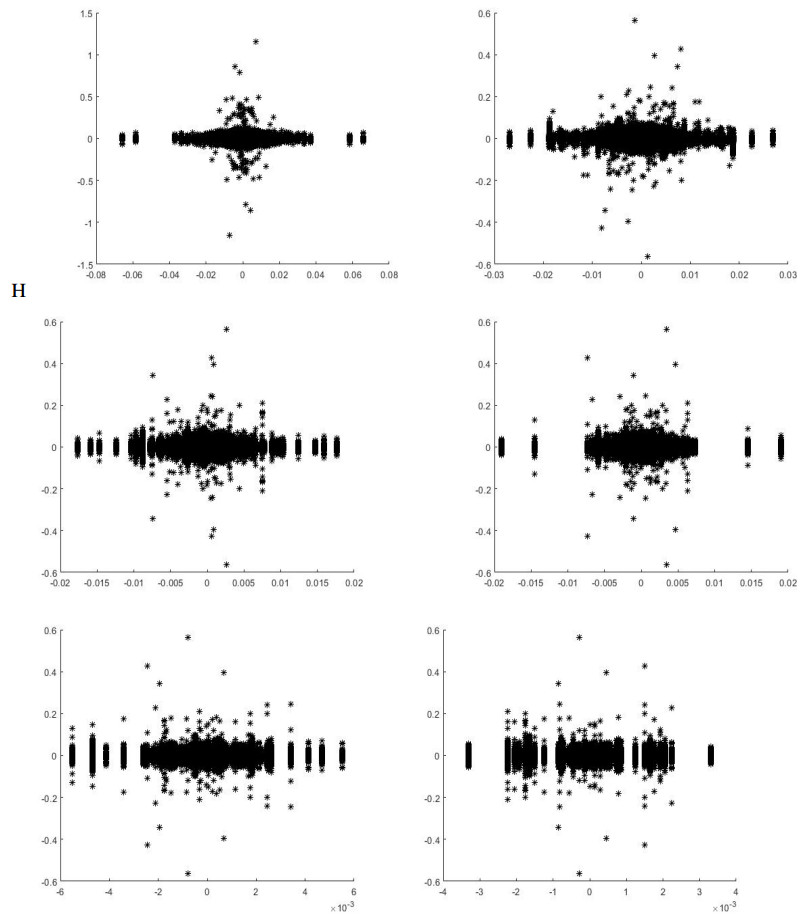
Systematic risk estimation is widely applied by investors and managers in order to predict risks in the market. One of the most applied measures of risk is the so-called Capital Asset Pricing Model, shortly CAPM. It has been studied empirically focusing on the impact of return interval on the betas. This paper lies in this topic and attempts to estimate the CAPM at different time scales for GCC markets by adapting a wavelet method to examine the relationship between the return of the stock and its systematic risk at different time scales. The main novelty is by applying non-uniform intervals of time. Differently from existing literature, we use random ones. The proposed procedure is acted empirically on a sample corresponding to Saudi Tadawul market as the most important GCC representative market actively traded over the period January 01, 2013 to September 20, 2018, which is characterized by many political, economic and financial movements such as Qatar embargo, Yemen war, NEOM project, 2030 KSA vision and the Arab spring effects. The findings in the present work may be good basis for understanding current and future GCC markets situation and may be thus a basis for investors' decisions in such markets.
Citation: Anouar Ben Mabrouk. Wavelet-based systematic risk estimation: application on GCC stock markets: the Saudi Arabia case[J]. Quantitative Finance and Economics, 2020, 4(4): 542-595. doi: 10.3934/QFE.2020026
| [1] | Daniel A. Barone, Ana C. Krieger . The Function of Sleep. AIMS Neuroscience, 2015, 2(2): 71-90. doi: 10.3934/Neuroscience.2015.2.71 |
| [2] | Eduardo Mercado III . Relating Cortical Wave Dynamics to Learning and Remembering. AIMS Neuroscience, 2014, 1(3): 185-209. doi: 10.3934/Neuroscience.2014.3.185 |
| [3] | Alexis M. Chambers, Jessica D. Payne . The Influence of Sleep on the Consolidation of Positive Emotional Memories: Preliminary Evidence. AIMS Neuroscience, 2014, 1(1): 39-51. doi: 10.3934/Neuroscience.2014.1.39 |
| [4] | Samson Z Assefa, Montserrat Diaz-Abad, Emerson M Wickwire, Steven M Scharf . The Functions of Sleep. AIMS Neuroscience, 2015, 2(3): 155-171. doi: 10.3934/Neuroscience.2015.3.155 |
| [5] | Marie Vandekerckhove, Yu-lin Wang . Emotion, emotion regulation and sleep: An intimate relationship. AIMS Neuroscience, 2018, 5(1): 1-17. doi: 10.3934/Neuroscience.2018.1.1 |
| [6] | Mark R. Zielinski, James T. McKenna, Robert W. McCarley . Functions and Mechanisms of Sleep. AIMS Neuroscience, 2016, 3(1): 67-104. doi: 10.3934/Neuroscience.2016.1.67 |
| [7] | Kimberly A Cote, Catherine E Milner, Tamara A Speth . Altered Sleep Mechanisms following Traumatic Brain Injury and Relation to Waking Function. AIMS Neuroscience, 2015, 2(4): 203-228. doi: 10.3934/Neuroscience.2015.4.203 |
| [8] | Carolyn E. Jones, Miranda M. Lim . Phasic Sleep Events Shape Cognitive Function after Traumatic Brain Injury: Implications for the Study of Sleep in Neurodevelopmental Disorders. AIMS Neuroscience, 2016, 3(2): 232-236. doi: 10.3934/Neuroscience.2016.2.232 |
| [9] | Ritwik Das, Artur Luczak . Epileptic seizures and link to memory processes. AIMS Neuroscience, 2022, 9(1): 114-127. doi: 10.3934/Neuroscience.2022007 |
| [10] | Shun Sakuma, Yuko Mizuno-Matsumoto, Yoshi Nishitani, Shinichi Tamura . Learning Times Required to Identify the Stimulated Position and Shortening of Propagation Path by Hebb’s Rule in Neural Network. AIMS Neuroscience, 2017, 4(4): 238-253. doi: 10.3934/Neuroscience.2017.4.238 |
Systematic risk estimation is widely applied by investors and managers in order to predict risks in the market. One of the most applied measures of risk is the so-called Capital Asset Pricing Model, shortly CAPM. It has been studied empirically focusing on the impact of return interval on the betas. This paper lies in this topic and attempts to estimate the CAPM at different time scales for GCC markets by adapting a wavelet method to examine the relationship between the return of the stock and its systematic risk at different time scales. The main novelty is by applying non-uniform intervals of time. Differently from existing literature, we use random ones. The proposed procedure is acted empirically on a sample corresponding to Saudi Tadawul market as the most important GCC representative market actively traded over the period January 01, 2013 to September 20, 2018, which is characterized by many political, economic and financial movements such as Qatar embargo, Yemen war, NEOM project, 2030 KSA vision and the Arab spring effects. The findings in the present work may be good basis for understanding current and future GCC markets situation and may be thus a basis for investors' decisions in such markets.
Originally likened to a wax tablet by Plato and Aristotle [1], memory has been the muse of philosophers and scientists alike. The discovery of the synapse separating individual neurons by early neuroscientists at the turn of the 20th century, and the subsequent proposal that increased activity at these synapses strengthens the connection between neurons [2] were pivotal ideas for expanding our understanding of memory beyond a philosophical concept or observed behavior to a complex process located in the brain. Today, we know that synaptic plasticity is important for memory formation, but there is still much to learn about how memory is formed at the neural level. As Cadonic and Albensi [3] point out, neural oscillations and receptors implicated in brain plasticity may hold the key to unlocking the mystery of how we form memories. Another essential player in this relationship, increasingly receiving support for its role in various memory processes, is sleep.
The ability to form new memories relies on synaptic plasticity, whereby high-frequency activation of a neural pathway increases the strength of later transmission across the affected synapses, a process termed long term potentiation (LTP) [4,5]. Amino-acid receptors have been implicated in the process of LTP, including the N-methyl-D-aspartate (NMDA) receptor and the alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptor. During LTP, the NMDA receptor is first activated when both the Mg2+ blocking its channel is removed by depolarization, and co-agonists glutamate and glycine are bound to its receptors [5,6]. This allows Ca2+ to flow into the cell, which initiates a cascade of events, including the activation of synaptic proteins and the phosphorylation and transfer of AMPA receptors into the synapse, culminating in the strengthening of excitatory postsynaptic currents [7]. LTP consists of two phases: the initial phase may last for hours, while the longer maintenance phase lasts for weeks [8]. Many studies connect LTP to the formation of new memories. For instance, an increase in the phosphorylation and number of AMPA receptors at the synapse has been found for LTP established via high-frequency stimulation in vitro, as well as during inhibitory avoidance learning in rats [9]. Blocking LTP has also been shown to block new learning [10].
Additionally, glutamatergic receptors like the NMDA receptor are important for the generation of neural oscillations, which have also been connected to memory formation. The rhythmic activity produced by the brain can be measured by electrophysiological measures such as the electroencephalogram (EEG), which has been used to delineate several frequency bands implicated in memory processes, including theta (4-7 Hz), alpha (7-14 Hz), beta (15-30 Hz), and gamma (30-100 Hz) frequencies [11,12]. Previous studies have found that NMDA antagonists block NMDA function and dramatically reduce electrically stimulated oscillations in the 4-9 Hz range [13]. Further, removal of the NMDA receptor in genetically altered mice decreases the power of theta and increases the power of gamma oscillations measured in the hippocampus compared to controls. This alteration negatively impacts spatial working-memory of these mice during a maze-learning task [14], which is in line with the proposal that such oscillations underlie varying types of memory formation and may represent transfer of information between brain regions [15]. In humans, changes in alpha, beta, gamma, and theta power are associated with memory for previously studied words [16], with encoding and retrieval strategies playing a role in the direction of such relationships [12].
While the review by Cadonic and Albensi [3] nicely outlines this relationship between glutamatergic receptors and oscillatory activity, pointing toward an important and fruitful area for future research on memory formation, further information may be provided by including sleep in this discussion, especially for the maintenance of already initiated LTP. Sleep is characterized by the alternation of dynamic stages, including non-rapid eye movement sleep (NREM), composed of stages 1, 2, and 3/4 or slow wave sleep (SWS), and rapid eye movement sleep (REM), each of which can be distinguished by unique physiological, chemical, and electrical properties [17,18]. Most important for the discussion here, NREM sleep, and particularly SWS, is characterized by slow oscillatory EEG patterns in the 0.5-4 Hz range, representing the alternation between up states (i.e., neural activity) and down states (i.e., neural silence), and punctuated by high-frequency bursts known as sleep spindles (12-15 Hz) in the cortex. These spindles are temporally correlated with sharp-wave ripples (150-250 Hz) in the hippocampus [19]. Conversely, REM sleep is characterized by theta EEG activity, which is synchronized with gamma oscillations in the hippocampus [18,20].
One of the most substantiated functions of sleep is its role in brain plasticity supporting both declarative and non-declarative memory processing [19]. The purported mechanism by which sleep has such a ubiquitous effect on memory is the reactivation and consolidation of recently potentiated synapses [21,22]. The Synaptic Re-entry Reinforcement hypothesis [21,22,23] proposes that the longevity of brain plasticity requires a reactivation of NMDA receptors following the initial learning event. Indeed, without proper synaptic stimulation and protein synthesis, LTP does not last beyond the several hours of its initial phase [24,25]. Thus, following the initiation of LTP during a learning event, potentiated synapses require reactivation to maintain synaptic plasticity. Sleep offers an optimum context for such reactivation to occur, given that activation of glutamate receptors and generation of neural oscillations are hallmarks of sleep processes [19].
Evidence for the reactivation of glutamatergic receptor mechanisms at potentiated synapses during sleep is abundant. Following monocular deprivation learning, whereby one eye is occluded to prevent input of visual information, neural activity becomes biased toward the region of visual cortex corresponding to the non-deprived eye, and this is further enhanced by a period of sleep. This effect is promoted by reactivation of potentiated pathways during sleep, as blocking NMDA receptors or the synthesis of related downstream proteins during a post-learning sleep period negates the consolidation of this plasticity. Further, phosphorylation of AMPA receptors is found only in animals allowed to sleep after monocular deprivation [22]. In humans, the blockade of NMDA or AMPA receptors during an eight-hour period of nocturnal, post-learning sleep likewise abolishes typical sleep-dependent improvements in visual discrimination learning [21]. Moreover, sleep deprivation, which is known to impair learning, has been shown to negatively alter glutamate receptor activation and protein cascades necessary for LTP [19,26]. This work highlights the importance of sleep for glutamatergic receptor signaling that maintains plasticity initiated during a waking period, without which learning would be impeded.
As others have pointed out, the oscillatory activity that occurs during sleep is similar to that supporting induction of LTP [19]. This may make sleep-related oscillations ideal for the reactivation of potentiated synapses. NREM sleep in particular has been the focus of many studies on this matter [27,28,29]. Indeed, some studies have found that under anesthesia or during waking, synapses previously strengthened by LTP are more likely to generate slow oscillatory up states and spindle activity [30,31]. Further, there is a reciprocal relationship between spindle activity and glutamatergic receptors, such that NMDA inputs mediate spindle activity [13], and stimulation of spindle activity in the cortex influences NMDA receptors and Ca2+, leading to the induction of LTP [29]. Consistently, neural electrical stimulation during in vitro imitations of the electrophysiological states that characterize SWS, including the combination of up states and down states, facilitates LTP and is blocked by NMDA or AMPA antagonists [28]. In vivo applications of slow oscillatory activity (0.75 Hz) during NREM sleep following the learning of word-pairs not only increases SWS, slow oscillatory power, and slow spindle power, but also results in better memory for the learned word-pairs compared to a control condition. Such effects are specific to slow oscillations, as theta frequency stimulation does not produce these results [27]. This evidence suggests a close relationship between the electrical activity of NREM sleep and activation of glutamate receptors that contribute to learning, providing evidence for the role of NREM sleep in reactivation and maintenance of LTP.
REM sleep is also implicated in LTP reprocessing. During REM sleep, but not NREM sleep, proteins necessary for the long-term maintenance of already established LTP, such as cAMP (cyclic adenosine monophosphate) and CREB (cAMP-response element binding protein), are activated at higher levels compared to a period of wakefulness, a process not found in mice genetically altered to lack memory consolidation abilities [19,32]. Antagonists acting on specific subunits of NMDA receptors also increase gamma activity during REM sleep [33]. This finding is interesting given that theta and gamma oscillations, which are synchronized within regions of the hippocampus during REM sleep with periodic bursts of synchrony across areas, are proposed to play a role in memory processing and transfer during sleep [20]. Theta and gamma oscillations are also temporally associated between hemispheres during REM, which may be particularly important for neural processes given that recovery sleep following REM deprivation produces a rebound and intensification of gamma synchronization between the left and right frontopolar and dorsolateral regions of the frontal lobe [34]. Importantly, stimulation in the gamma range across the corpus callosum initiates a short-lived potentiation that is blocked with NMDA antagonists, implicating this oscillatory connection between hemispheres during REM sleep as another mechanism for the sleep-associated maintenance of plasticity in neural circuits [34,35].
As some of the most interesting evidence for neuronal replay during sleep, hippocampal place cells that were activated during spatial learning during a waking period are sequentially reactivated on a compressed timescale during subsequent NREM and on a timescale comparable to learning during subsequent REM [36,37]. Important to the discussion here, replay is associated with sharp wave ripples in the hippocampus during NREM sleep, and theta activity during REM sleep. Given that these electrical markers of sleep have been correlated with memory improvements [38,39], such replay is proposed to reflect a mechanism of memory consolidation through an exchange in communication between the hippocampus and cortex [18,40].
It is important to note that another hypothesis regarding sleep and neural plasticity, the Synaptic Homeostasis Hypothesis [41,42], proposes that sleep generally downscales the potentiation of synapses that has accumulated over a period wakefulness, thus maintaining neural stability [42]. This hypothesis puts particular emphasis on SWS and slow oscillations, which are seen to increase after waking, and decrease after sleep [42]. While seemingly at odds with the studies discussed thus far, which show that sleep processes help maintain LTP, others have suggested that synaptic downscaling and maintenance may both occur during SWS, depending on how the plasticity relates to different types of learning [22], and how it may develop in different neural pathways that differ in their requirements for plasticity [28].
As Cadonic and Albensi [3] point out, abnormal NMDA receptor signaling also likely plays a role in pathology, such that hypo- and hyperactivation has been associated with Schizophrenia and Alzheimer’s disease, respectively. Not surprisingly, evidence additionally indicates oscillatory activity, and sleep, in the etiology of such diseases. Wamsley and colleagues [43] have demonstrated that Schizophrenia patients display a reduction in the number and density of sleep spindles during a period of sleep following motor task learning, which is associated with a reduction in motor performance following sleep compared to controls. High-frequency (> 20 Hz) oscillatory power is also increased during sleep in Schizophrenic and depressed patients compared to healthy individuals [44]. While these studies do not clarify exact causal relationships, they reaffirm the role of sleep in brain plasticity and cognitive functioning, and may provide clues to developing treatments.
Ample research points toward sleep as a crucial contributor to neural plasticity and learning. The dynamic neurophysiology of sleep provides an optimum milieu for the reactivation of recently potentiated synapses and transfer of information to cortical stores, with alterations of this environment linked to pathology. Given the foregoing discussion, Cadonic and Albensi [3] are accurate to champion the further investigation of plasticity-related receptors and oscillations in future memory research, but the addition of sleep to this endeavor will greatly expand the progress we can make to ultimately understanding how we remember and learn.
All the authors declare to have no conflict of interest.
| [1] | Aktan B, Ozturk M, Rhaeim N, et al. (2009) Wavelet-Based Systematic Risk Estimation An Application on Istanbul Stock Exchange. Int Res J Financ Econ 23: 34-45. |
| [2] | Arfaoui S, Rezgui I, Mabrouk AB (2017) Wavelet Analysis On The Sphere, Spheroidal Wavelets, Degryuter, Degruyter, 2017, ISBN 978-3-11-048188-4. |
| [3] | Arfaoui S, Mabrouk AB, Cattani C (2020) New type of Gegenbauer-Hermite monogenic polynomials and associated Clifford wavelets. J Math Imaging Vision 62: 73-97. |
| [4] | Arfaoui S, Mabrouk AB, Cattani C (2020) New type of Gegenbauer-Jacobi-Hermite monogenic polynomials and associated continuous Clifford wavelet transform. Acta Applicandea Math. https://doi.org/10.1007/s10440-020-00322-0. |
| [5] | Aydogan K (1989) Portfolio Theory, Capital Market Board of Turkey Research Report: Ankara. |
| [6] | Banz RW (1981) The relationship between return and market value of common stock. J Financ Econ 9: 3-18. |
| [7] | Basu S (1977) The relationship between earnings' yield, market value and return for NYSE common stocks-further evidence. J Financ 32: 663-681. |
| [8] | Mabrouk AB, Mohamed MLB, Omrani K (2008) Numerical solutions for PDEs modeling binary alloy-solidification dynamics, In: Proceedings of 2007 International Symposium on Nonlinear Dynamics, J Phys, conference series 96 (2008) 012067. |
| [9] | Mabrouk AB, Kortass H, Ammou SB (2008) Wavelet Estimators for Long Memory in Stock Markets. Int J Theor Appl Financ 12: 297-317. |
| [10] | Mabrouk AB, Kahloul I, Hallara SE (2010) Wavelet-Based Prediction for Governance, Diversification and Value Creation Variables. Int Res J Financ Econ 60: 15-28. |
| [11] | Mabrouk AB, Abdallah NB, Hamrita ME (2011) A wavelet method coupled with quasi self similar stochastic processes for time series approximation. Int J Wavelets Multiresolution Inf Process 9: 685-711. |
| [12] | Mabrouk AB, Zaafrane O (2013) Wavelet Fuzzy Hybrid Model For Physico Financial Signals. J Appl Stat 40: 1453-1463. |
| [13] | Mabrouk AB, Rabbouch B, Saadaoui F (2015) A wavelet based methodology for predicting transmembrane segments, In: Poster Session, The International Conference of Engineering Sciences for Biology and Medecine, 1-3 May 2015, Monastir, Tunisie. |
| [14] | Black F, Jensen MC, Scholes M(1972) The capital asset pricing model: Some empirical tests, in Jensen MC (ed), Studies in the theory of Capital, New York: Praeger, 1-54. |
| [15] | Black F (1972) Capital Market Equilibrium with Restricted Borrowing. J Bus 45: 444-455. |
| [16] | Breeden D (1979) An Intertemporal Asset Pricing Model with Stochastic Consumption and Investment Opportunities. J Financ Econ 73: 265-296. |
| [17] | Brennan MJ (1973) Taxes, market valuation and corporate financial policy. Natl tax J 23: 417-427. |
| [18] | Chae J, Yang C (2008) Which idiosyncratic factors can explain the pricing errors from asset pricing models in the Korean stock market? Asia-Pasific J Financ Stud 37: 297-342. |
| [19] | Chan LK, Lakonishok J (1993) Are the reports of beta's death premature? J Portf Manage 19: 51-62. |
| [20] | Cifter A, Ozun A (2007) Multiscale systematic risk: An application on ISE 30, MPRA Paper 2484, University Library of Munich: Germany. |
| [21] | Cifter A, Ozun A (2008) A signal processing model for time series analysis: The effect of international F/X markets on domestic currencies using wavelet networks. Int Rev Electr Eng 3: 580-591. |
| [22] | Cohen K, Hawawin G, Mayer S, et al. (1986) The Microstructure of Securities Markets, Prentice-Hall: Sydney. |
| [23] | Conlon T, Crane M, Ruskin HJ (2008) Wavelet multiscale analysis for hedge funds: Scaling and strategies. Phys A 387: 5197-5204. |
| [24] | Daubechies I (1992) Ten Lectures on Wavelets, Society for Industrial and Applied Mathematics, Philadelphia. |
| [25] | Desmoulins-Lebeault F (2003) Distribution of Returns and the CAPM Empirical Problems. Post-Print halshs-00165099, HAL. |
| [26] | DiSario R, Saroglu H, McCarthy J, et al. (2008) Long memory in the volatility of an emerging equity market: The case of Turkey. Int Mark Inst Money 18: 305-312. |
| [27] | Fama EF (1970) Efficient capital markets: A review of theory and empirical work. J Financ 25: 383-417. |
| [28] | Fama E, French K (1992) The Cross-section of Expected Stock Returns. J Financ 47: 427-465. |
| [29] | Fama E, French K (1993) Common risk factors in returns on stocks and bonds. J Financ Econ 33: 3-56. |
| [30] | Fama E, French KR (1996) The CAPM is Wanted, Dead or Alive. J Financ 51: 1947-1958. |
| [31] | Fama E, French KR (2004) The Capital Asset Pricing Model: Theory and Evidence. J Econo Perspect 18: 25-46. |
| [32] | Fama E, French KR (2006) The Value Premium and the CAPM. J Financ 61: 2163-2185. |
| [33] | Fama E, MacBeth J (1973) Risk, return and equilibrium: Empirical tests. J Polit Econ 81: 607-636. |
| [34] | Fernandez V (2006) The CAPM and value at risk at different time-scales. Int Rev Financ Anal 15: 203-219. |
| [35] | Friend L, Landskroner Y, Losq E (1976) The demand for risky assets and uncertain inflation. J Financ 31: 1287-1297. |
| [36] | Galagedera DUA (2007) A review of capital asset pricing models. Managerial Financ 33: 821-832. |
| [37] | Gençay R, Selçuk F, Whitcher B (2002) An Introduction to Wavelets and Other Filtering Methods in Finance and Economics, Academic Press, San Diego. |
| [38] | Gençay R, Whitcher B, Selçuk F (2003) Systematic Risk and Time Scales. Quant Financ 3: 108-116. |
| [39] | Gençay R, Whitcher B, Selçuk F (2005) Multiscale systematic risk. J Inte Money Financ 24: 55-70. |
| [40] | Gibbons MR (1982) Multivariate tests of financial models: A new approach. J Financ Econ 10: 3-27. |
| [41] | Gursoy CT, Rejepova G (2007) Test of capital asset pricing model in Turkey. J Dogus Univ 8: 47-58. |
| [42] | Handa P, Kothari SP, Wasley C (1989) The relation between the return interval and beta: Implications for size-effect. J Financ Econ 23: 79-100. |
| [43] | Handa P, Kothari SP, Wasley C (1993) Sensitivity of multivariate tests of the CAPM to the return measurement interval. J Financ 48: 1543-1551. |
| [44] | Ho YW, Strange R, Piesse J (2000) CAPM anomalies and the pricing of equity: Evidence from the Hong Kong market. Appl Econ 32: 1629-1636. |
| [45] | Hubbard BB (1998) The world according to wavelets: The story of a mathematical technique in the making, 2e, Ak Peters Ltd., MA. |
| [46] | Mahmoud IMM, Ben Mabrouk A, Hashim MHA (2016) Wavelet multifractal models for transmembrane proteins' series. Int J Wavelets Multires Inf Process 14: 36. |
| [47] | In F, Kim S (2006) The hedge ratio and the empirical relationship between the stock and futures markets: A new approach using wavelet analysis. J Bus 79: 799-820. |
| [48] | In F, Kim S (2007) A note on the relationship between Fama-French risk factors and innovations of ICAPM state variables. Financ Res Lett 4: 165-171. |
| [49] | In F, Kim S, Marisetty V, et al. (2008) Analysing the performance of managed funds using the wavelet multiscaling method. Rev Quant Financ Accounting 31: 55-70. |
| [50] | Karan MB, Karadagli E (2001) Risk return and market equilibrium in Istanbul stock exchange: The test of the capital asset pricing model. J Econ Administrative Sci 19: 165-177. |
| [51] | Kishor NK, Marfatia HA (2013) The time-varying response of foreign stock markets to US monetary policy surprises: Evidence from the Federal funds futures market. J Int Financ Mark Inst Money 24: 1-24. |
| [52] | Kothari S, Shanken J (1998) On defense of beta, J. Stern, and D. Chew, Jr. (Eds.), The Revolution in Corporate Finance, 3e, Blackwell Publishers Inc., 52-57. |
| [53] | Levhari D, Levy H (1977) The Capital Asset Pricing Model and the Investment Horizon. Rev Econ Stat 59: 92-104. |
| [54] | Lévy H (1978) Equilibrium in an imperfect market: a constraint on the number of securities in the portfolio. Am Econ Rev 68: 643-658. |
| [55] | Lintner J (1965a) Security Prices and Maximal Gaines from Diversification. J Financ 20: 587-615. |
| [56] | Lintner J (1965b) The Valuation of Risk Assets and the Selection of Risky Investments in Stock Portfolios and Capital Budgets. Rev Econ Stat 47: 13-37. |
| [57] | Litzenberger RH, Ramaswamy K (1979) The effect of personal taxes and dividends on capital asset prices: Theory and empirical evidence. J Financ Econ 7: 163-195. |
| [58] | Magni CA (2007a) Project selection and equivalent CAPM-based investment criteria. Appl Financ Econ Lett 3: 165-168. |
| [59] | Magni CA (2007b) Project valuation and investment decisions: CAPM versus arbitrage. Appl Financ Econ Lett 3: 137-140. |
| [60] | Marfatia HA (2014) Impact of uncertainty on high frequency response of the US stock markets to the Fed's policy surprises. Q Rev Econ Financ 54: 382-392. |
| [61] | Marfatia HA (2015) Monetary policy's time-varying impact on the US bond markets: Role of financial stress and risks. North Am J Econ Financ 34: 103-123. |
| [62] | Marfatia HA (2017a) A fresh look at integration of risks in the international stock markets: A wavelet approach. Rev Financ Econ 34: 33-49. |
| [63] | Marfatia HA (2017b) Wavelet Linkages of Global Housing Markets and macroeconomy. Available at SSRN 3169424. |
| [64] | Marfatia HA (2020) Investors' Risk Perceptions in the US and Global Stock Market Integration. Res Int Bus Financ 52: 101169. |
| [65] | Markowitz H (1952) Portfolio Selection. J Financ 7: 77-91. |
| [66] | Merton RC (1973) An Intertermporal Capital Asset Pricing Model. Econometrica: J Econometric Society 41: 867-887. |
| [67] | Mossin I (1966) Equilibrium in a Capital Asset Market. Econometrica: J Econometric Society 34: 768-783. |
| [68] | Perold A (2004) The Capital Asset Pricing Model. J Econ Perspect 18: 3-24. |
| [69] | Percival DB, Walden AT (2000) Wavelet methods for time series analysis, Camridge University Press, NY. |
| [70] | Rhaiem R, Ammou SB, Mabrouk AB (2007a) Estimation of the systematic risk at different time scales: Application to French stock market. Int J Appl Econ Financ 1: 79-87. |
| [71] | Rhaiem R, Ammou SB, Mabrouk AB (2007b) Wavelet estimation of systematic risk at different time scales, Application to French stock markets. Int J Appl Econ Financ 1: 113-119. |
| [72] | Roll R (1977) A critique of the asset pricing theory's tests Part I: On past and potential testability of the theory. J Financ Econ 4: 129-176. |
| [73] | Selcuk F (2005) Wavelets: A new analysis method (in Turkish). Bilkent J 3: 12-14. |
| [74] | Sharkasi A, Crane M, Ruskin HJ, et al.(2006) The reaction of stock markets to crashes and events: A comparison study between emerging and mature markets using wavelet transforms. Phys A 368: 511-521. |
| [75] | Sharpe WF (1964) Capital asset prices: A theory of market equilibrium under conditions of risk. J Financ 19: 425-442. |
| [76] | Sharpe WF (1970a) Computer-Assisted Economics. J Financ Quant Anal, 353-366. |
| [77] | Sharpe WF (1970b) Stock market price behavior. A discussion. J Financ 25: 418-420. |
| [78] | Sharpe WF (1970c) Portfolio theory and capital markets, McGraw-Hill College. |
| [79] | Soltani S (2002) On the use of the wavelet decomposition for time series prediction. Neurocomput 48: 267-277. |
| [80] | Soltani S, Modarres R, Eslamian SS (2007) The use of time series modeling for the determination of rainfall climates of Iran. Int J Climatol 27: 819-829. |
| [81] | Vasichek AA, McQuown JA (1972) Le modèle de marché efficace. Analyse financière, 15, 1973, traduit de "The effecient market model". Financ Anal J. |
| [82] | Xiong X, Zhang X, Zhang W, et al. (2005) Wavelet-based beta estimation of China stock market, In: Proceedings of 4th International Conference on Machine Learning and Cybernetic, Guangzhou. IEEE: 0-7803-9091-1. |
| [83] | Yamada H (2005) Wavelet-based beta estimation and Japanese industrial stock prices. Appl Econ Lett 12: 85-88. |
| [84] | Zemni M, Jallouli M, Mabrouk AB, et al. (2019a) Explicit Haar-Schauder multiwavelet filters and algorithms. Part II: Relative entropy-based estimation for optimal modeling of biomedical signals. Int J Wavelets Multiresolution Inf Process 17: 1950038. |
| [85] | Zemni M, Jallouli M, Mabrouk AB, et al. (2019b) ECG Signal Processing with Haar-Schauder Multiwavelet, In: Proceedings of the 9th International Conference on Information Systems and Technologies—Icist 2019. |
 QFE-04-04-026-s001.pdf QFE-04-04-026-s001.pdf |
 |
Figures(3) / Tables(9)

Anouar Ben Mabrouk. Wavelet-based systematic risk estimation: application on GCC stock markets: the Saudi Arabia case[J]. Quantitative Finance and Economics, 2020, 4(4): 542-595. doi: 10.3934/QFE.2020026







 DownLoad:
DownLoad: