Citation: Valentina Bashkatova, Athineos Philippu. Role of nitric oxide in psychostimulant-induced neurotoxicity[J]. AIMS Neuroscience, 2019, 6(3): 191-203. doi: 10.3934/Neuroscience.2019.3.191

| [1] |
Dawson TM, Snyder SH (1994) Gases as biological messengers: nitric oxide and carbon monoxide in the brain. J Neurosci 14: 5147–5159. doi: 10.1523/JNEUROSCI.14-09-05147.1994

|
| [2] |
Moncada S, Higgs EA (1991) Endogenous nitric oxide: physiology, pathology and clinical relevance. Eur J Clin Invest 21: 361–374. doi: 10.1111/j.1365-2362.1991.tb01383.x
|
| [3] |
Philippu A (2016) Nitric Oxide: A universal modulator of brain function. Curr Med Chem 23: 2643–2652. doi: 10.2174/0929867323666160627120408
|
| [4] |
Garthwaite J (2019) NO as a multimodal transmitter in the brain: discovery and current status. Br J Pharmacol 176: 197–211. doi: 10.1111/bph.14532
|
| [5] | Ignarro LJ (1990) Nitric oxide. A novel signal transduction mechanism for transcellular communication. Hypertension 16: 477–483. |
| [6] |
Möller MN, Cuevasanta E, Orrico F, et al. (2019) Diffusion and transport of reactive species across cell membranes. Adv Exp Med Biol 1127: 3–19. doi: 10.1007/978-3-030-11488-6_1
|
| [7] |
Li H, Förstermann U (2000) Nitric oxide in the pathogenesis of vascular disease. J Pathol 190: 244–254. doi: 10.1002/(SICI)1096-9896(200002)190:3<244::AID-PATH575>3.0.CO;2-8
|
| [8] |
Prast H, Philippu A (2001) Nitric oxide as a modulator of neuronal function. Prog Neurobiol 64: 51–68. doi: 10.1016/S0301-0082(00)00044-7
|
| [9] |
Mukherjee P, Cinelli MA, Kang S, et al. (2014) Development of nitric oxide synthase inhibitors for neurodegeneration and neuropathic pain. Chem Soc Rev 43: 6814–6838. doi: 10.1039/C3CS60467E
|
| [10] |
Capannolo M, Ciccarelli C, Molteni R, et al. (2014) Nitric oxide synthase inhibition reverts muscarinic receptor down-regulation induced by pilocarpine- and kainic acid-evoked seizures in rat fronto-parietal cortex. Epilepsy Res 108: 11–19. doi: 10.1016/j.eplepsyres.2013.10.011
|
| [11] |
Watanabe S, Kumazaki S, Yamamoto S, et al. (2018) Non-alcoholic steatohepatitis aggravates nitric oxide synthase inhibition-induced arteriosclerosis in SHRSP5/Dmcr rat model. Int J Exp Pathol 99: 282–294. doi: 10.1111/iep.12301
|
| [12] |
Woodard A, Barbery B, Wilkinson R, et al. (2019) The role of neuronal nitric oxide and its pathways in the protection and recovery from neurotoxin-induced de novo hypokinetic motor behaviors in the embryonic zebrafish (Danio rerio). AIMS Neuroscience 6: 25–42. doi: 10.3934/Neuroscience.2019.1.25
|
| [13] |
Bredt DS, Snyder SH (1994) Nitric oxide: A physiologic messenger molecule. Annu Rev Biochem 63: 175–195. doi: 10.1146/annurev.bi.63.070194.001135
|
| [14] |
Sorokin A (2016) Nitric oxide synthase and cyclooxygenase pathways: A complex interplay in cellular signaling. Curr Med Chem 23: 2559–2578. doi: 10.2174/0929867323666160729105312
|
| [15] |
Balke JE, Zhang L, Percival JM (2019) Neuronal nitric oxide synthase (nNOS) splice variant function: Insights into nitric oxide signaling from skeletal muscle. Nitric Oxide 82: 35–47. doi: 10.1016/j.niox.2018.11.004
|
| [16] |
Teixeira-Gomes A, Costa VM, Feio-Azevedo R, et al. (2015) The neurotoxicity of amphetamines during the adolescent period. Int J Dev Neurosci 41: 44–62. doi: 10.1016/j.ijdevneu.2014.12.001
|
| [17] | Jones SR, Joseph JD, Barak LS, et al. (1999) Dopamine neuronal transport kinetics and effects of amphetamine. J Neurochem 73: 2406–2414. |
| [18] | Gibb JW, Johnson M, Hanson GR (1990) Neurochemical basis of neurotoxicity. Neurotoxicol 11: 317–321. |
| [19] |
Sulzer D, Rayport S (1990) Amphetamine and other psychostimulants reduce pH gradients in midbrain dopaminergic neurons and chromaffin granules: a mechanism of action. Neuron 5: 797–808. doi: 10.1016/0896-6273(90)90339-H
|
| [20] |
Siciliano CA, Calipari ES, Ferris MJ, et al. (2014) Biphasic mechanisms of amphetamine action at the dopamine terminal. J Neurosci 34: 5575–5582. doi: 10.1523/JNEUROSCI.4050-13.2014
|
| [21] |
O'Dell SJ, Weihmuller FB, Marshall JF (1991) Multiple methamphetamine injections induce marked increases in extracellular striatal dopamine which correlate with subsequent neurotoxicity. Brain Res 564: 256–260. doi: 10.1016/0006-8993(91)91461-9
|
| [22] |
Nash JF, Yamamoto BK (1993) Effect of D-amphetamine on the extracellular concentrations of glutamate and dopamine in iprindole-treated rats. Brain Res 627: 1–8. doi: 10.1016/0006-8993(93)90741-5
|
| [23] |
Hussain RJ, Taraschenko OD, Glick SD (2008) Effects of nicotine, methamphetamine and cocaine on extracellular levels of acetylcholine in the interpeduncular nucleus of rats. Neurosci Lett 440: 270–274. doi: 10.1016/j.neulet.2008.06.001
|
| [24] |
Mabrouk OS, Semaan DZ, Mikelman S, et al. (2014) Amphetamine stimulates movement through thalamocortical glutamate release. J Neurochem 128: 152–161. doi: 10.1111/jnc.12378
|
| [25] |
Abekawa T, Ohmori T, Honda M, et al. (2001) Effect of low doses of L-NAME on methamphetamine-induced dopaminergic depletion in the rat striatum. J Neural Transm 108: 1219–1230. doi: 10.1007/s007020100000
|
| [26] |
Pereira FC, Macedo TR, Imam SZ, et al. (2004) Lack of hydroxyl radical generation upon central administration of methamphetamine in rat caudate nucleus: A microdialysis study. Neurotox Res 6: 149–152. doi: 10.1007/BF03033217
|
| [27] |
Shenouda SK, Varner KJ, Carvalho F, et al. (2009) Metabolites of MDMA induce oxidative stress and contractile dysfunction in adult rat left ventricular myocytes. Cardiovasc Toxicol 9: 30–38. doi: 10.1007/s12012-009-9034-6
|
| [28] |
Salum C, Schmidt F, Michel PP, et al. (2016) Signaling mechanisms in the Nitric Oxide donor-and amphetamine-induced dopamine release in mesencephalic primary cultured neurons. Neurotox Res 29: 92–104. doi: 10.1007/s12640-015-9562-8
|
| [29] |
Acikgoz O, Gonenc S, Kayatekin BM, et al. (2000) The effects of single dose of methamphetamine on lipid peroxidation levels in the rat striatum and prefrontal cortex. Eur Neuropsychopharmacol 10: 415–418. doi: 10.1016/S0924-977X(00)00103-6
|
| [30] |
Wan FJ, Lin HC, Huang KL, et al. (2000) Systemic administration of d-amphetamine induces long-lasting oxidative stress in the rat striatum. Life Sci 66: 205–212. doi: 10.1016/S0024-3205(00)00458-6
|
| [31] | Raevskii KS, Bashkatova VG, Narkevich VB, et al. (1998) Nitric oxide in the rat cerebral cortex in seizure models: potential ways of pharmacological modulation. Ross Fiziol Zh Im I M Sechenova 84: 1093–1099. |
| [32] | Bashkatova VG, Vitskova GIu, Narkevich VB, et al. (1999) The effect of anticonvulsants on the nitric oxide content and level of lipid peroxidation in the brain of rats in model seizure states. Eksp Klin Farmakol 62: 11–14. |
| [33] | Fadiukova OE, Alekseev AA, Bashkatova VG, et al. (2001) Semax prevents elevation of nitric oxide generation caused by incomplete global ischemia in the rat brain. Eksp Klin Farmakol 64: 31–34. |
| [34] |
Zheng Y, Laverty R (1998) Role of brain nitric oxide in (+/−)3,4-methylenedioxymethamphetamine (MDMA)-induced neurotoxicity in rats. Brain Res 795: 257–263. doi: 10.1016/S0006-8993(98)00313-8
|
| [35] |
Itzhak Y, Martin JL, Ail SF (2000) nNOS inhibitors attenuate methamphetamine-induced dopaminergic neurotoxicity but not hyperthermia in mice. Neuroreport 11: 2943–2946. doi: 10.1097/00001756-200009110-00022
|
| [36] |
Liu YP, Tung CS, Lin PJ, et al. (2011) Role of nitric oxide in amphetamine-induced sensitization of schedule-induced polydipsic rats. Psychopharmacology (Berl) 218: 599–608. doi: 10.1007/s00213-011-2354-9
|
| [37] |
Morales-Medina JC, Mejorada A, Romero-Curiel A, et al. (2008) Neonatal administration of N-omega-nitro-L-arginine induces permanent decrease in NO levels and hyperresponsiveness to locomotor activity by D-amphetamine in postpubertal rats. Neuropharmacology 55: 1313–1320. doi: 10.1016/j.neuropharm.2008.08.019
|
| [38] |
De Silva DJ, French SJ, Cheung NY, et al. (2005) Rat brain serotonin neurones that express neuronal nitric oxide synthase have increased sensitivity to the substituted amphetamine serotonin toxins 3,4-methylenedioxymethamphetamine and p-chloroamphetamine. Neuroscience 134: 1363–1375. doi: 10.1016/j.neuroscience.2005.05.016
|
| [39] |
Wang JQ, Lau YS (2001) Dose-related alteration in nitric oxide synthase mRNA expression induced by amphetamine and the full D1 dopamine receptor agonist SKF-82958 in mouse striatum. Neurosci Lett 311: 5–8. doi: 10.1016/S0304-3940(01)02128-0
|
| [40] |
Tocharus J, Chongthammakun S, Govitrapong P (2008) Melatonin inhibits amphetamine-induced nitric oxide synthase mRNA overexpression in microglial cell lines. Neurosci Lett 439: 134–137. doi: 10.1016/j.neulet.2008.05.036
|
| [41] |
Kleschyov AL, Sedov KR, Mordvintcev PI, et al. (1994) Biotransformation of sodium nitroprusside into dinitrosyl iron complexes in tissue of ascites tumors of mice. Biochem Biophys Res Commun 202: 168–173. doi: 10.1006/bbrc.1994.1908
|
| [42] |
Vanin AF, Huisman A, van Faassen EE (2002) Iron dithiocarbamate as spin trap for nitric oxide detection: pitfalls and successes. Methods Enzymol 359: 27–42. doi: 10.1016/S0076-6879(02)59169-2
|
| [43] |
Hogg N (2010) Detection of nitric oxide by electron paramagnetic resonance spectroscopy. Free Radic Biol Med 49: 122–129. doi: 10.1016/j.freeradbiomed.2010.03.009
|
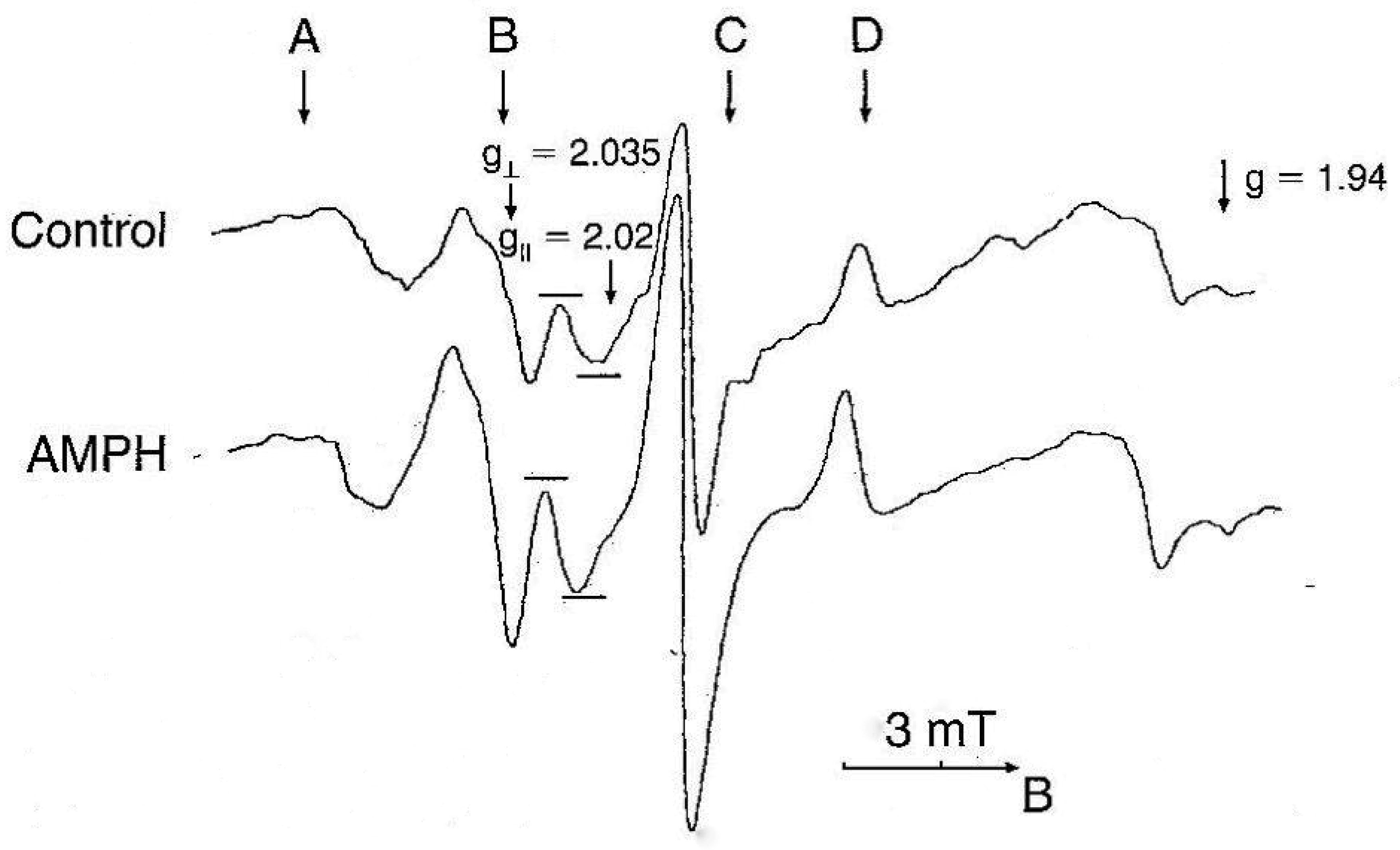
| [44] | Bashkatova VG, Mikoian VD, Kosacheva ES, et al. (1996) Direct determination of nitric oxide in rat brain during various types of seizures using ESR. Dokl Akad Nauk 348: 119–121. |
| [45] |
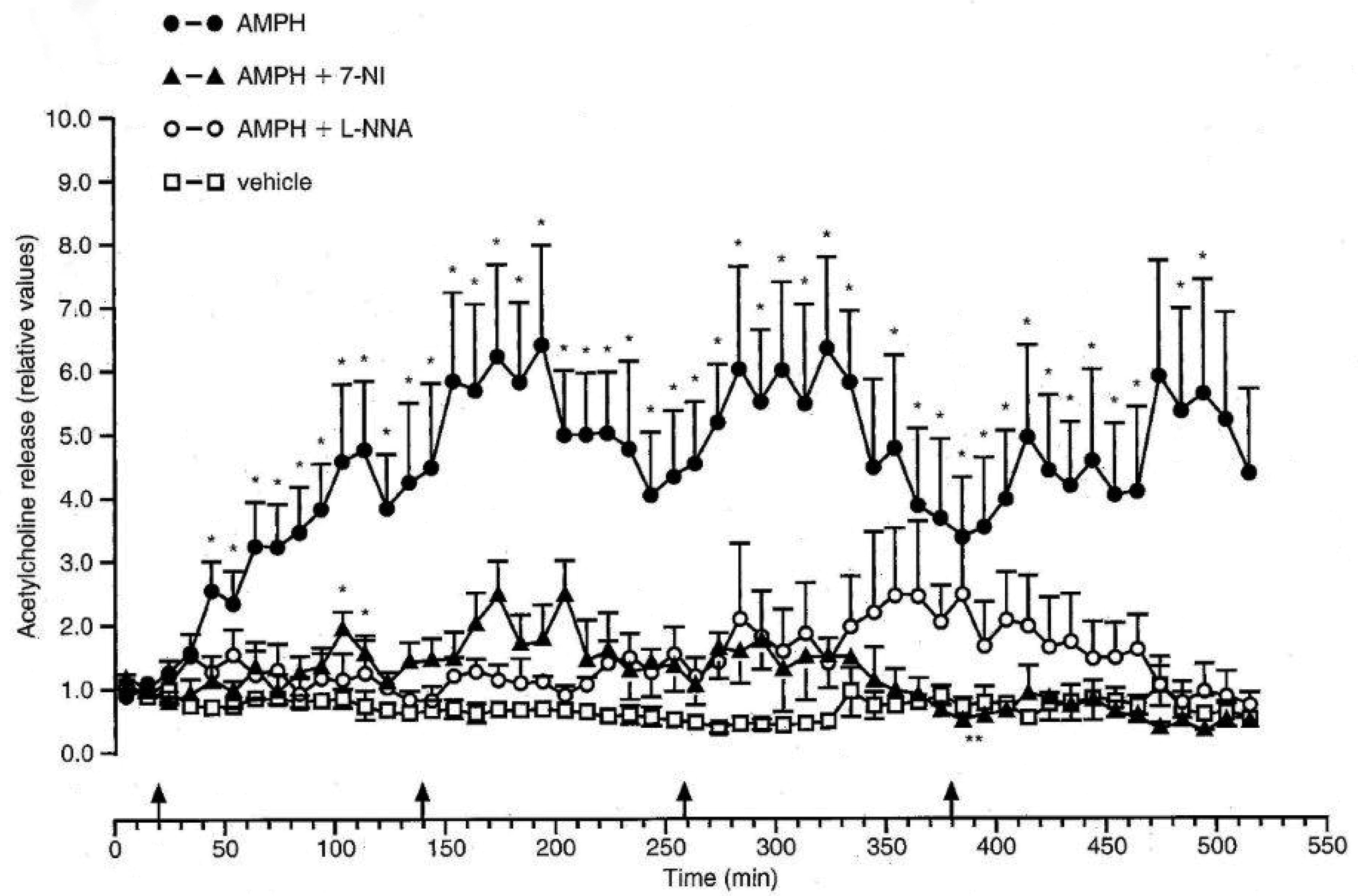
Bashkatova V, Kraus M, Prast H, et al. (1999) Influence of NOS inhibitors on changes in ACH release and NO level in the brain elicited by amphetamine neurotoxicity. Neuroreport 10: 3155–3158. doi: 10.1097/00001756-199910190-00006
|
| [46] |
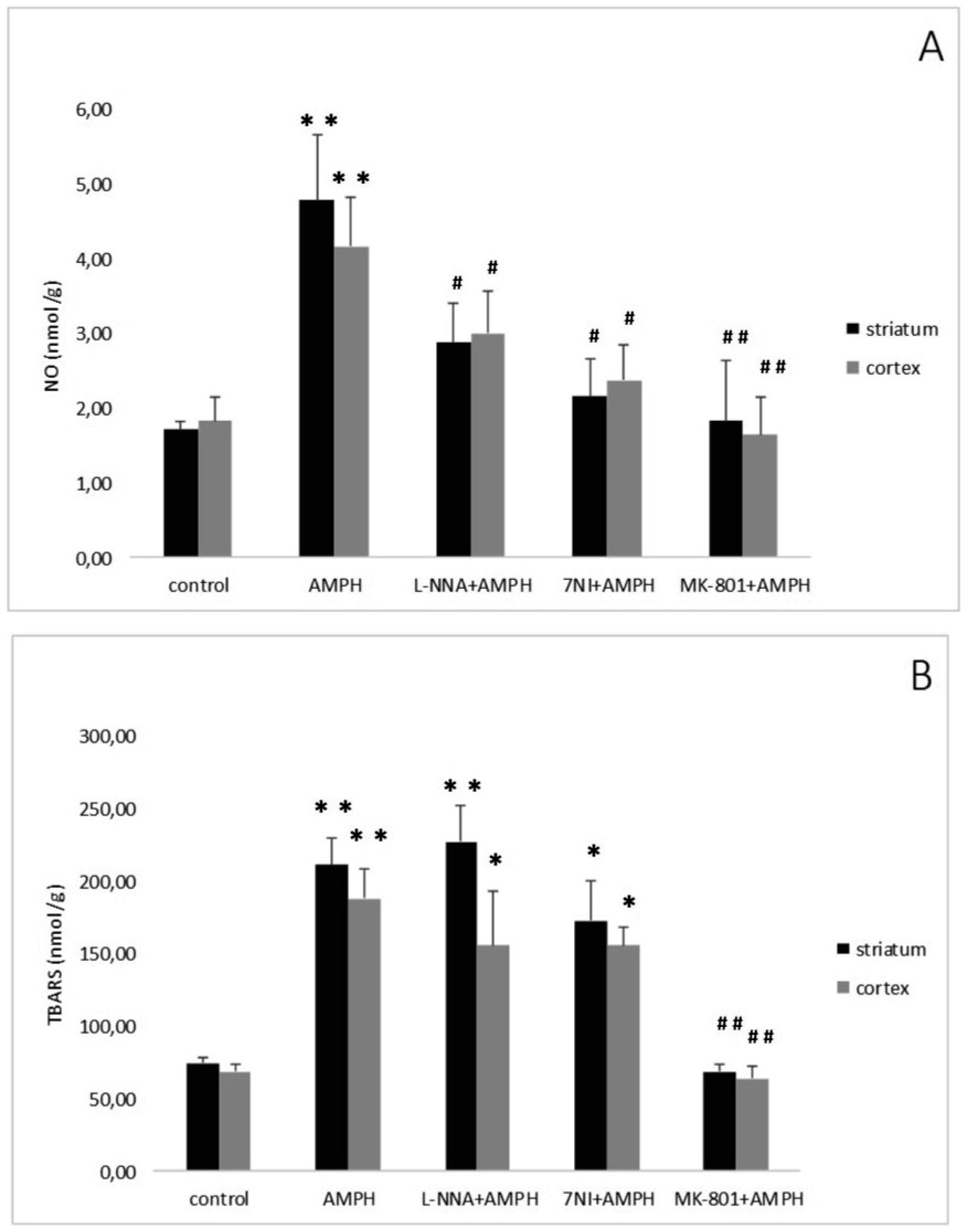
Bashkatova V, Kraus MM, Vanin A, et al. (2005) 7-Nitroindazole, nNOS inhibitor, attenuates amphetamine-induced amino acid release and nitric oxide generation but not lipid peroxidation in the rat brain. J Neural Transm 112: 779–788. doi: 10.1007/s00702-004-0224-x
|
| [47] |
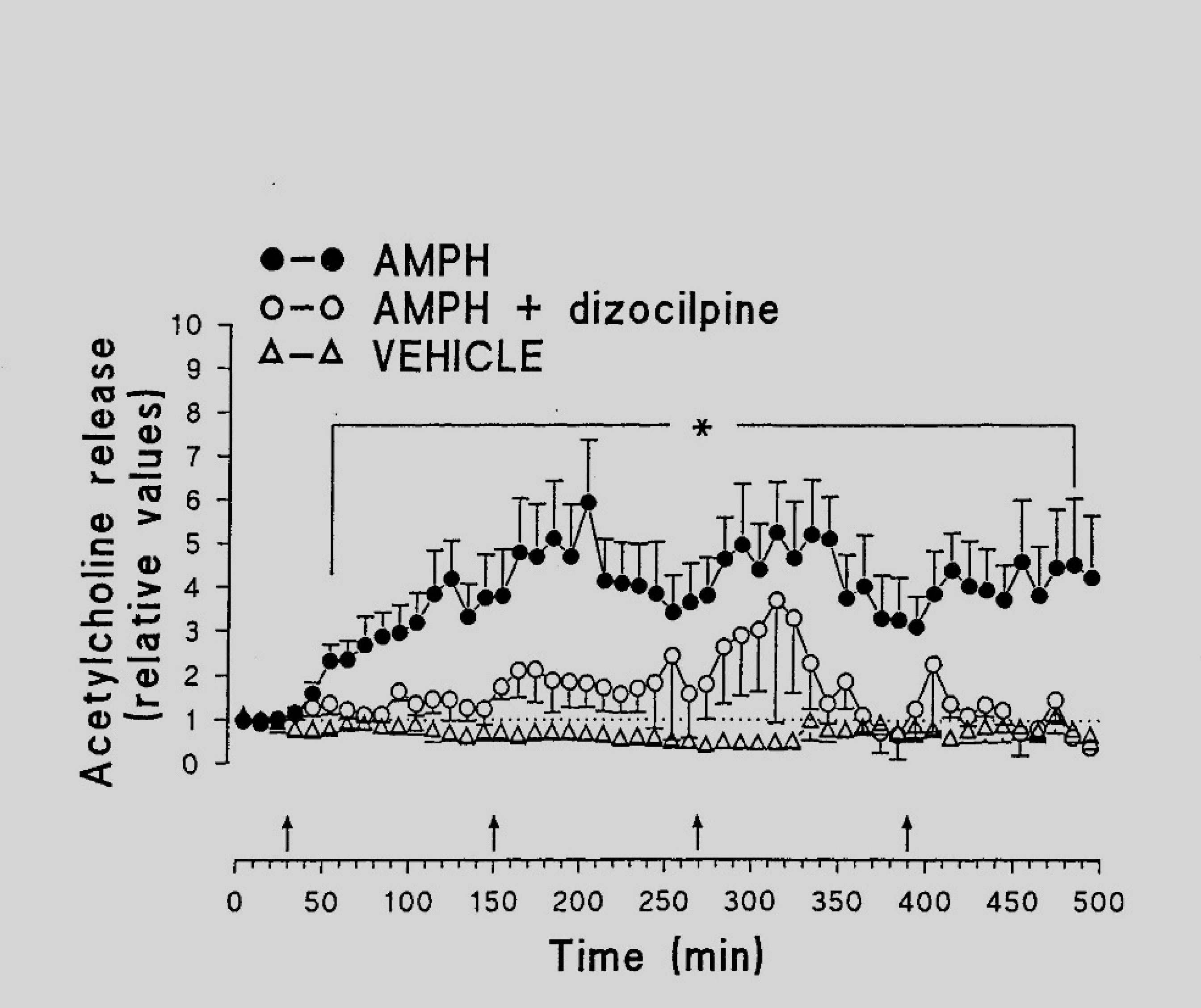
Kraus MM, Bashkatova V, Vanin A, et al. (2002) Dizocilpine inhibits amphetamine-induced formation of nitric oxide and amphetamine-induced release of amino acids and acetylcholine in the rat brain. Neurochem Res 27: 229–235. doi: 10.1023/A:1014836621717
|
| [48] |
Wan FJ, Tung CS, Shiah IS, et al. (2006) Effects of alpha-phenyl-N-tert-butyl nitrone and N- acetylcysteine on hydroxyl radical formation and dopamine depletion in the rat striatum produced by d-amphetamine. Eur Neuropsychopharmacol 16: 147–153. doi: 10.1016/j.euroneuro.2005.07.002
|
| [49] |
Kita T, Miyazaki I, Asanuma M, et al. (2009) Dopamine-induced behavioral changes and oxidative stress in methamphetamine-induced neurotoxicity. Int Rev Neurobiol 88: 43–64. doi: 10.1016/S0074-7742(09)88003-3
|
| [50] | Dawson TM, Dawson VL, Snyder SH (1994) Molecular mechanisms of nitric oxide actions in the brain. Ann N Y Acad Sci 738: 76–85. |
| [51] |
Li J, Baud O, Vartanian T, et al. (2005) Peroxynitrite generated by inducible nitric oxide synthase and NADPH oxidase mediates microglial toxicity to oligodendrocytes. Proc Natl Acad Sci U S A 102: 9936–9941. doi: 10.1073/pnas.0502552102
|
| [52] |
Ali SF, Imam SZ, Itzhak Y (2005) Role of peroxynitrite in methamphetamine-induced dopaminergic neurodegeneration and neuroprotection by antioxidants and selective NOS inhibitors. Ann N Y Acad Sci 1053: 97–98. doi: 10.1196/annals.1344.053
|
| [53] |
Ohkawa H, Ohishi N, Yagi K (1979) Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem 95: 351–358. doi: 10.1016/0003-2697(79)90738-3
|
| [54] |
Bashkatova V, Vitskova G, Narkevich V, et al. (2000) Nitric oxide content measured by ESR-spectroscopy in the rat brain is increased during pentylenetetrazole-induced seizures. J Mol Neurosci 14: 183–190. doi: 10.1385/JMN:14:3:183
|
| [55] |
Klyueva YYu, Chepurnov SA, Chepurnova NE, et al. (2001) Role of nitric oxide and lipid peroxidation in mechanisms of febrile convulsions in wistar rat pups. Bull Exp Biol Med 131: 47–49. doi: 10.1023/A:1017530612936
|
| [56] | Prast H, Fischer H, Werner E, et al. (1995) Nitric oxide modulates the release of acetylcholine in the ventral striatum of the freely moving rat. Naunyn-Schmiedebergґs Arch Pharmacol 352: 67–73. |
| [57] |
Issy AC, Dos-Santos-Pereira M, Pedrazzi JFC, et al. (2018) The role of striatum and prefrontal cortex in the prevention of amphetamine-induced schizophrenia-like effects mediated by nitric oxide compounds. Prog Neuropsychopharmacol Biol Psychiatry 86: 353–362. doi: 10.1016/j.pnpbp.2018.03.015
|
| [58] |
Salum C, Guimarães FS, Brandão ML, et al. (2006) Dopamine and nitric oxide interaction on the modulation of prepulse inhibition of the acoustic startle response in the Wistar rat. Psychopharmacology (Berl) 185: 133–141. doi: 10.1007/s00213-005-0277-z
|
| [59] |
Sonsalla PK, Nicklas WJ, Heikkila RE (1989) Role for excitatory amino acids in methamphetamine-induced nigrostriatal dopaminergic toxicity. Science 243: 398–400. doi: 10.1126/science.2563176
|
| [60] |
Tata DA, Yamamoto BK (2007) Interactions between methamphetamine and environmentalstress: role of oxidative stress, glutamate and mitochondrial dysfunction. Addiction 102: 49–60. doi: 10.1111/j.1360-0443.2007.01770.x
|
| [61] |
Li MH, Underhill SM, Reed C, et al. (2017) Amphetamine and methamphetamine increaseNMDAR-GluN2B synaptic currents in midbrain dopamine neurons. Neuropsychopharmacology 42: 1539–1547. doi: 10.1038/npp.2016.278
|
| [62] |
Haj-Mirzaian A, Amiri S, Amini-Khoei H, et al. (2018) Involvement of NO/NMDA-R pathwayin the behavioral despair induced by amphetamine withdrawal. Brain Res Bull 139: 81–90. doi: 10.1016/j.brainresbull.2018.02.001
|
| [63] |
Erenberg G (2005) The relationship between tourette syndrome, attention deficit hyperactivitydisorder, and stimulant medication: a critical review. Semin Pediatr Neurol 12: 217–221. doi: 10.1016/j.spen.2005.12.003
|
| [64] | Jain R, Jain S, Montano CB (2017) Addressing diagnosis and treatment gaps in adults withattention-deficit/hyperactivity disorder. Prim Care Companion CNS Disord 19: pii: 17nr02153. |
| [65] |
Quilty LC, Allen TA, Davis C, et al. (2019) A randomized comparison of long actingmethylphenidate and cognitive behavioral therapy in the treatment of binge eating disorder. Psychiatry Res 273: 467–474. doi: 10.1016/j.psychres.2019.01.066
|
| [66] | Novoselov IA, Cherepov AB, Raevskii KS, et al. (2002) Locomotor activity and expression ofc-Fos protein in the brain of C57BL and Balb/c mice: effects of D-amphetamine and sydnocarb. Eksp Klin Farmakol 65: 18–21. |
| [67] |
Gruner JA, Mathiasen JR, Flood DG, et al. (2011) Characterization of pharmacological andwake-promoting properties of the dopaminergic stimulant sydnocarb in rats. J Pharmacol ExpTher 337: 380–390. doi: 10.1124/jpet.111.178947
|
| [68] |
Gainetdinov RR, Sotnikova TD, Grekhova TV, et al. (1997) Effects of a psychostimulant drugsydnocarb on rat brain dopaminergic transmission in vivo. Eur J Pharmacol 340: 53–58. doi: 10.1016/S0014-2999(97)01407-6
|
| [69] | Afanas'ev II, Anderzhanova EA, Kudrin VS, et al. (2001) Effects of amphetamine andsydnocarb on dopamine release and free radical generation in rat striatum. Pharmacol BiochemBehav 200169: 653–658. |
| [70] | Bashkatova V, Mathieu-Kia AM, Durand C, et al. (2002) Neurochemical changes and neurotoxic effects of an acute treatment with sydnocarb, a novel psychostimulant: comparison with D-amphetamine. Ann N Y Acad Sci 965: 180–192. |
| [71] |
Feier G, Valvassori SS, Lopes-Borges J (2012) Behavioral changes and brain energy metabolism dysfunction in rats treated with methamphetamine or dextroamphetamine. Neurosci Lett 530: 75–79. doi: 10.1016/j.neulet.2012.09.039
|
| [72] | Witkin JM, Savtchenko N, Mashkovsky M, et al. (1999) Behavioral, toxic, and neurochemical effects of sydnocarb, a novel psychomotor stimulant: comparisons with methamphetamine. J Pharmacol Exp Ther 288: 1298–1310. |
| [73] |
Anderzhanova EA, Afanas'ev II, Kudrin VS, et al. (2000) Effect of d-amphetamine and sydnocarb on the extracellular level of dopamine, 3,4-dihydroxyphenylacetic acid, and hydroxyl radicals generation in rat striatum. Ann N Y Acad Sci 914: 137–145. doi: 10.1111/j.1749-6632.2000.tb05191.x
|
| [74] |
Chen N, Li J, Li D, et al. (2014) Chronic exposure to perfluorooctane sulfonate induces behavior defects and neurotoxicity through oxidative damages, in vivo and in vitro. PLoS One 9: e113453. doi: 10.1371/journal.pone.0113453
|
| [75] |
Zhang LP, Wang QS, Guo X, et al. (2007) Time-dependent changes of lipid peroxidation and antioxidative status in nerve tissues of hens treated with tri-ortho-cresyl phosphate (TOCP). Toxicology 239: 45–52. doi: 10.1016/j.tox.2007.06.091
|
Figures(4)

Valentina Bashkatova, Athineos Philippu. Role of nitric oxide in psychostimulant-induced neurotoxicity[J]. AIMS Neuroscience, 2019, 6(3): 191-203. doi: 10.3934/Neuroscience.2019.3.191







 DownLoad:
DownLoad: