Citation: Constance Jeffery. Intracellular proteins moonlighting as bacterial adhesion factors[J]. AIMS Microbiology, 2018, 4(2): 362-376. doi: 10.3934/microbiol.2018.2.362

| [1] |
Jeffery CJ (1999) Moonlighting proteins. Trends Biochem Sci 24: 8–11. doi: 10.1016/S0968-0004(98)01335-8

|
| [2] | Henderson B, Martin A (2011) Bacterial virulence in the moonlight: multitasking bacterial moonlighting proteins are virulence determinants in infectious disease. Infect Immun 79: 347691. |
| [3] | Henderson B, Martin A (2013) Bacterial moonlighting proteins and bacterial virulence. Curr Top Microbiol 358: 155–213. |
| [4] |
Jeffery CJ (2009) Moonlighting proteins-an update. Mol Biosyst 5: 345–350. doi: 10.1039/b900658n
|
| [5] |
Kainulainen V, Korhonen TK (2014) Dancing to another tune-adhesive moonlighting proteins in bacteria. Biology 3: 178–204. doi: 10.3390/biology3010178
|
| [6] |
Pancholi V, Fischetti VA (1992) A major surface protein on group A streptococci is a glyceraldehyde-3-phosphate-dehydrogenase with multiple binding activity. J Exp Med 176: 415–426. doi: 10.1084/jem.176.2.415
|
| [7] |
Mani M, Chen C, Amblee V, et al. (2015) MoonProt: a database for proteins that are known to moonlight. Nucleic Acids Res 43: D277–D282. doi: 10.1093/nar/gku954
|
| [8] |
Agarwal S, Kulshreshtha P, Bambah MD, et al. (2008) Alpha-enolase binds to human plasminogen on the surface of Bacillus anthracis. BBA-Proteins Proteom 1784: 986–994. doi: 10.1016/j.bbapap.2008.03.017
|
| [9] |
Bao S, Guo X, Yu S, et al. (2014) Mycoplasma synoviae enolase is a plasminogen/fibronectin binding protein. BMC Vet Res 10: 223. doi: 10.1186/s12917-014-0223-6
|
| [10] |
Boleij A, Laarakkers CM, Gloerich J, et al. (2011) Surface-affinity profiling to identify host-pathogen interactions. Infect Immun 79: 4777–4783. doi: 10.1128/IAI.05572-11
|
| [11] |
Candela M, Biagi E, Centanni M, et al. (2009) Bifidobacterial enolase, a cell surface receptor for human plasminogen involved in the interaction with the host. Microbiology 155: 3294–3303. doi: 10.1099/mic.0.028795-0
|
| [12] |
Jones MN, Holt RG (2007) Cloning and characterization of an alpha-enolase of the oral pathogen Streptococcus mutans that binds human plasminogen. Biochem Bioph Res Co 364: 924–929. doi: 10.1016/j.bbrc.2007.10.098
|
| [13] |
Vanegas G, Quiñones W, Carrasco-López C, et al. (2007) Enolase as a plasminogen binding protein in Leishmania mexicana. Parasitol Res 101: 1511–1516. doi: 10.1007/s00436-007-0668-7
|
| [14] | Antikainen J, Kuparinen V, Lähteenmäki K, et al. (2007) Enolases from Gram-positive bacterial pathogens and commensal lactobacilli share functional similarity in virulence-associated traits FEMS Immunol Med Mic 51: 526–534. |
| [15] |
Esgleas M, Li Y, Hancock MA, et al. (2008) Isolation and characterization of alpha-enolase, a novel fibronectin-binding protein from Streptococcus suis. Microbiology 154: 2668–2679. doi: 10.1099/mic.0.2008/017145-0
|
| [16] |
Floden AM, Watt JA, Brissette CA (2011) Borrelia burgdorferi enolase is a surface-exposed plasminogen binding protein. PLoS One 6: e27502. doi: 10.1371/journal.pone.0027502
|
| [17] | Fulde M, Rohde M, Polok A, et al. (2013) Cooperative plasminogen recruitment to the surface of Streptococcus canis via M protein and enolase enhances bacterial survival. MBio 4: e00629-12. |
| [18] |
Kesimer M, Kilic N, Mehrotra R, et al. (2009) Identification of salivary mucin MUC7 binding proteins from Streptococcus gordonii. BMC Microbiol 9: 163. doi: 10.1186/1471-2180-9-163
|
| [19] |
Kinnby B, Booth NA, Svensater G (2008) Plasminogen binding by oral streptococci from dental plaque and inflammatory lesions. Microbiology 154: 924–931. doi: 10.1099/mic.0.2007/013235-0
|
| [20] |
Knaust A, Weber MV, Hammerschmidt S, et al. (2007) Cytosolic proteins contribute to surface plasminogen recruitment of Neisseria meningitidis. J Bacteriol 189: 3246–3255. doi: 10.1128/JB.01966-06
|
| [21] |
Carneiro CR, Postol E, Nomizo R, et al. (2004) Identification of enolase as a laminin-binding protein on the surface of Staphylococcus aureus. Microbes Infect 6: 604–608. doi: 10.1016/j.micinf.2004.02.003
|
| [22] |
Sha J, Erova TE, Alyea RA, et al. (2009) Surface-expressed enolase contributes to the pathogenesis of clinical isolate SSU of Aeromonas hydrophila. J Bacteriol 191: 3095–3107. doi: 10.1128/JB.00005-09
|
| [23] |
Castaldo C, Vastano V, Siciliano RA, et al. (2009) Surface displaced alfa-enolase of Lactobacillus plantarum is a fibronectin binding protein. Microb Cell Fact 8: 14. doi: 10.1186/1475-2859-8-14
|
| [24] |
Yavlovich A, Rechnitzer H, Rottem S (2007) Alpha-enolase resides on the cell surface of Mycoplasma fermentans and binds plasminogen. Infect Immun 75: 5716–5719. doi: 10.1128/IAI.01049-07
|
| [25] |
Tunio SA, Oldfield NJ, Berry A, et al. (2010) The moonlighting protein fructose-1, 6-bisphosphate aldolase of Neisseria meningitidis: surface localization and role in host cell adhesion. Mol Microbiol 76: 605–615. doi: 10.1111/j.1365-2958.2010.07098.x
|
| [26] |
Blau K, Portnoi M, Shagan M, et al. (2007) Flamingo cadherin: a putative host receptor for Streptococcus pneumoniae. J Infect Dis 195: 1828–1837. doi: 10.1086/518038
|
| [27] |
Hennequin C, Porcheray F, Waligora-Dupriet A, et al. (2001) GroEL (Hsp60) of Clostridium difficile is involved in cell adherence. Microbiology 147: 87–96. doi: 10.1099/00221287-147-1-87
|
| [28] | Yamaguchi H, Osaki T, Kurihara N, et al. (1997) Heat-shock protein 60 homologue of Helicobacter pylori is associated with adhesion of H. pylori to human gastric epithelial cells. J Med Microbiol 46: 825–831. |
| [29] |
Wuppermann FN, Molleken K, Julien M, et al. (2008) Chlamydia pneumoniae GroEL1 protein is cell surface associated and required for infection of HEp-2 cells. J Bacteriol 190: 3757–3767. doi: 10.1128/JB.01638-07
|
| [30] | Garduno RA, Garduno E, Hoffman PS (1998) Surface-associated hsp60 chaperonin of Legionella pneumophila mediates invasion in a HeLa cell model. Infect Immun 66: 4602–4610. |
| [31] |
Jagadeesan B, Koo OK, Kim KP, et al. (2010) LAP, an alcohol acetaldehyde dehydrogenase enzyme in Listeria, promotes bacterial adhesion to enterocyte-like Caco-2 cells only in pathogenic species. Microbiology 156: 2782–2795. doi: 10.1099/mic.0.036509-0
|
| [32] |
Wampler JL, Kim KP, Jaradat Z, et al. (2004) Heat shock protein 60 acts as a receptor for the Listeria adhesion protein in Caco-2 cells. Infect Immun 72: 931–936. doi: 10.1128/IAI.72.2.931-936.2004
|
| [33] |
Jin H, Song YP, Boel G, et al. (2005) Group A streptococcal surface GAPDH, SDH, recognizes uPAR/CD87 as its receptor on the human pharyngeal cell and mediates bacterial adherence to host cells. J Mol Biol 350: 27–41. doi: 10.1016/j.jmb.2005.04.063
|
| [34] | Frisk AC, Ison CA, Lagergard T (1998) GroEL heat shock protein of Haemophilus ducreyi: association with cell surface and capacity to bind to eukaryotic cells. Infect Immun 66: 1252–1257. |
| [35] | Pantzar M, Teneberg S, Lagergard T (2006) Binding of Haemophilus ducreyi to carbohydrate receptors is mediated by the 58.5-kDa GroEL heat shock protein. Microbes Infect 8: 2452–2458. |
| [36] |
Dallo SF, Kannan TR, Blaylock MW, et al. (2002) Elongation factor Tu and E1 beta subunit of pyruvate dehydrogenase complex act as fibronectin binding proteins in Mycoplasma pneumoniae. Mol Microbiol 46: 1041–1051. doi: 10.1046/j.1365-2958.2002.03207.x
|
| [37] |
Heilmann C, Hartleib J, Hussain MS, et al. (2005) The multifunctional Staphylococcus aureus autolysin aaa mediates adherence to immobilized fibrinogen and fibronectin. Infect Immun 73: 4793–4802. doi: 10.1128/IAI.73.8.4793-4802.2005
|
| [38] | Kinhikar AG, Vargas D, Li H, et al. (2006) Mycobacterium tuberculosis malate synthase is a laminin-binding adhesin. Mol Microbiol 60: 999–1013. |
| [39] |
Jung CJ, Zheng QH, Shieh YH, et al. (2009) Streptococcus mutans autolysin AtlA is a fibronectin-binding protein and contributes to bacterial survival in the bloodstream and virulence for infective endocarditis. Mol Microbiol 74: 888–902. doi: 10.1111/j.1365-2958.2009.06903.x
|
| [40] |
Allignet J, England P, Old I, et al. (2002) Several regions of the repeat domain of the Staphylococcus caprae autolysin, AtlC, are involved in fibronectin binding. FEMS Microbiol Lett 213: 193–197. doi: 10.1111/j.1574-6968.2002.tb11305.x
|
| [41] |
Alvarez RA, Blaylock MW, Baseman JB (2003) Surface localized glyceraldehyde-3-phosphate dehydrogenase of Mycoplasma genitalium binds mucin. Mol Microbiol 48: 1417–1425. doi: 10.1046/j.1365-2958.2003.03518.x
|
| [42] | Ensgraber M, Loos M (1992) A 66-kilodalton heat shock protein of Salmonella typhimurium is responsible for binding of the bacterium to intestinal mucus. Infect Immun 60: 3072–3078. |
| [43] |
Kunert A, Losse J, Gruszin C, et al. (2007) Immune evasion of the human pathogen Pseudomonas aeruginosa: elongation factor Tuf is a factor H and plasminogen binding protein. J Immunol 179: 2979–2988. doi: 10.4049/jimmunol.179.5.2979
|
| [44] |
Raymond BB, Djordjevic S (2015) Exploitation of plasmin(ogen) by bacterial pathogens of veterinary significance. Vet Microbiol 178: 1–13. doi: 10.1016/j.vetmic.2015.04.008
|
| [45] | Collen D, Verstraete M (1975) Molecular biology of human plasminogen II Metabolism in physiological and some pathological conditions in man. Thromb Diath Haemorrh 34: 403–408. |
| [46] |
Dano K, Andreasen PA, Grondahl-Hansen J, et al. (1985) Plasminogen activators, tissue degradation, and cancer. Adv Cancer Res 44: 139–266. doi: 10.1016/S0065-230X(08)60028-7
|
| [47] |
Robinson MW, Buchtmann KA, Jenkins C, et al. (2013) MHJ_0125 is an M42 glutamyl aminopeptidase that moonlights as a multifunctional adhesin on the surface of Mycoplasma hyopneumoniae. Open Biol 3: 130017. doi: 10.1098/rsob.130017
|
| [48] |
Jarocki VM, Santos J, Tacchi JL, et al. (2015) MHJ_0461 is a multifunctional leucine aminopeptidase on the surface of Mycoplasma hyopneumoniae. Open Biol 5: 140175. doi: 10.1098/rsob.140175
|
| [49] | Modun B, Williams P (1999) The staphylococcal transferrin-binding protein is a cell wall glyceraldehyde-3-phosphate dehydrogenase. Infect Immun 67: 1086–1092. |
| [50] |
Kinoshita H, Uchida H, Kawai Y, et al. (2008) Cell surface Lactobacillus plantarum LA 318 glyceraldehyde-3-phosphate dehydrogenase (GAPDH) adheres to human colonic mucin. J Appl Microbiol 104: 1667–1674. doi: 10.1111/j.1365-2672.2007.03679.x
|
| [51] |
Granato D, Bergonzelli GE, Pridmore RD, et al. (2004) Cell surface-associated elongation factor Tu mediates the attachment of Lactobacillus johnsonii NCC533 (La1) to human intestinal cells and mucins. Infect Immun 72: 2160–2169. doi: 10.1128/IAI.72.4.2160-2169.2004
|
| [52] |
Bergonzelli GE, Granato D, Pridmore RD, et al. (2006) GroEL of Lactobacillus johnsonii La1 (NCC 533) is cell surface associated: potential role in interactions with the host and the gastric pathogen Helicobacter pylori. Infect Immun 74: 425–434. doi: 10.1128/IAI.74.1.425-434.2006
|
| [53] |
Patel DK, Shah KR, Pappachan A, et al. (2016) Cloning, expression and characterization of a mucin-binding GAPDH from Lactobacillus acidophilus. Int J Biol Macromol 91: 338–346. doi: 10.1016/j.ijbiomac.2016.04.041
|
| [54] |
Katakura Y, Sano R, Hashimoto T, et al. (2010) Lactic acid bacteria display on the cell surface cytosolic proteins that recognize yeast mannan. Appl Microbiol Biot 86: 319–326. doi: 10.1007/s00253-009-2295-y
|
| [55] |
Wang W, Jeffery CJ (2016) An analysis of surface proteomics results reveals novel candidates for intracellular/surface moonlighting proteins in bacteria. Mol Biosyst 12: 1420–1431. doi: 10.1039/C5MB00550G
|
| [56] |
Ehinger S, Schubert WD, Bergmann S, et al. (2004) Plasmin(ogen)-binding alpha-enolase from Streptococcus pneumoniae: crystal structure and evaluation of plasmin(ogen)-binding sites. J Mol Biol 343: 997–1005. doi: 10.1016/j.jmb.2004.08.088
|
| [57] | Green ER, Mecsas J (2016) Bacterial secretion systems: an overview. Microbiol Spectrum 4. |
| [58] |
Ebner P, Rinker J, Götz F (2016) Excretion of cytoplasmic proteins in Staphylococcus is most likely not due to cell lysis. Curr Genet 62: 19–23. doi: 10.1007/s00294-015-0504-z
|
| [59] |
Ebner P, Prax M, Nega M, et al. (2015) Excretion of cytoplasmic proteins (ECP) in Staphylococcus aureus. Mol Microbiol 97: 775–789. doi: 10.1111/mmi.13065
|
| [60] |
Yang CK, Ewis HE, Zhang X, et al. (2011) Nonclassical protein secretion by Bacillus subtilis in the stationary phase is not due to cell lysis. J Bacteriol 193: 5607–5615. doi: 10.1128/JB.05897-11
|
| [61] |
Yang CK, Zhang XZ, Lu CD, et al. (2014) An internal hydrophobic helical domain of Bacillus subtilis enolase is essential but not sufficient as a non-cleavable signal for its secretion. Biochem Bioph Res Co 446: 901–905. doi: 10.1016/j.bbrc.2014.03.032
|
| [62] |
Boël G, Pichereau V, Mijakovic I, et al. (2004) Is 2-phosphoglycerate-dependent automodification of bacterial enolases implicated in their export? J Mol Biol 337: 485–496. doi: 10.1016/j.jmb.2003.12.082
|
| [63] |
Amblee V, Jeffery CJ (2015) Physical features of intracellular proteins that moonlight on the cell surface. PLoS One 10: e0130575. doi: 10.1371/journal.pone.0130575
|
| [64] |
Widjaja M, Harvey KL, Hagemann L, et al. (2017) Elongation factor Tu is a multifunctional and processed moonlighting protein. Sci Rep 7: 11227. doi: 10.1038/s41598-017-10644-z
|
| [65] | Tacchi JL, Raymond BB, Haynes PA, et al. (2016) Post-translational processing targets functionally diverse proteins in Mycoplasma hyopneumoniae. Open Biol 6: 150210. |
| [66] | Navarre WW, Schneewind O (1999) Surface proteins of Gram positive bacteria and mechanisms of their targeting to the cell wall envelope. Microbiol Mol Biol R 63: 174–229. |
| [67] |
Schneewind O, Missiakas DM (2012) Protein secretion and surface display in Gram-positive bacteria. Philos T R Soc B 367: 1123–1139. doi: 10.1098/rstb.2011.0210
|
| [68] |
Scott JR, Barnett TC (2006) Surface proteins of Gram-positive bacteria and how they get there. Annu Rev Microbiol 60: 397–423. doi: 10.1146/annurev.micro.60.080805.142256
|
| [69] |
Desvaux M, Dumas E, Chafsey I, et al. (2006) Protein cell surface display in Gram-positive bacteria: from single protein to macromolecular protein structure. FEMS Microbiol Lett 256: 1–15. doi: 10.1111/j.1574-6968.2006.00122.x
|
| [70] |
Antikainen J, Kuparinen V, Lähteenmäki K, et al. (2007) pH-dependent association of enolase and glyceraldehyde-3-phosphate dehydrogenase of Lactobacillus crispatus with the cell wall and lipoteichoic acids. J Bacteriol 189: 4539–4543. doi: 10.1128/JB.00378-07
|
| [71] |
Sánchez B, Bressollier P, Urdaci MC (2008) Exported proteins in probiotic bacteria: adhesion to intestinal surfaces, host immunomodulation and molecular cross-talking with the host. FEMS Immunol Med Mic 54: 1–17. doi: 10.1111/j.1574-695X.2008.00454.x
|
| [72] |
Daubenspeck JM, Liu R, Dybvig K (2016) Rhamnose links moonlighting proteins to membrane phospholipid in mycoplasmas. PLoS One 11: e0162505. doi: 10.1371/journal.pone.0162505
|
| [73] |
Antikainen J, Kuparinen V, Lähteenmäki K, et al. (2007) pH-dependent association of enolase and glyceraldehyde-3-phosphate dehydrogenase of Lactobacillus crispatus with the cell wall and lipoteichoic acids. J Bacteriol 189: 4539–4543. doi: 10.1128/JB.00378-07
|
| [74] | Centers for Disease Control and Prevention, Antibiotic resistance threats in the United States, 2013. Available from: http://www.cdc.gov/drugresistance/threat-report-2013. |
| [75] | Ventola CL (2015) The antibiotic resistance crisis: part 1: causes and threats. Pharm Ther 40: 277–283. |
| [76] |
Matsuoka K, Kanai T (2015) The gut microbiota and inflammatory bowel disease. Semin Immunopathol 37: 47–55. doi: 10.1007/s00281-014-0454-4
|
| [77] | Dahlhamer JM, Zammitti EP, Ward BW, et al. (2016) Prevalence of inflammatory bowel disease among adults aged ≥18 years-United States, 2015. MMWR 65: 1166–1169. |
| [78] |
Kinnby B, Booth NA, Svensater G (2008) Plasminogen binding by oral streptococci from dental plaque and inflammatory lesions. Microbiology 154: 924–931. doi: 10.1099/mic.0.2007/013235-0
|
| [79] |
Heilmann C, Thumm G, Chhatwal GS, et al. (2003) Identification and characterization of a novel autolysin (Aae) with adhesive properties from Staphylococcus epidermidis. Microbiology 149: 2769–2778. doi: 10.1099/mic.0.26527-0
|
| [80] | Sjostrom I, Grondahl H, Falk G, et al. (1997) Purification and characterization of a plasminogen-binding protein from Haemophilus influenzae. Sequence determination reveals identity with aspartase. BBA-Biomembranes 1324: 182–190. |
| [81] |
Candela M, Bergmann S, Vici M, et al. (2007) Binding of human plasminogen to Bifidobacterium. J Bacteriol 189: 5929–5936. doi: 10.1128/JB.00159-07
|
| [82] |
Beckmann C, Waggoner JD, Harris TO, et al. (2002) Identification of novel adhesins from Group B streptococci by use of phage display reveals that C5a peptidase mediates fibronectin binding. Infect Immun 70: 2869–2876. doi: 10.1128/IAI.70.6.2869-2876.2002
|
| [83] |
Agarwal V, Kuchipudi A, Fulde M, et al. (2013) Streptococcus pneumoniae endopeptidase O (PepO) is a multifunctional plasminogen- and fibronectin-binding protein, facilitating evasion of innate immunity and invasion of host cells. J Biol Chem 288: 6849–6863. doi: 10.1074/jbc.M112.405530
|
| [84] |
Schreiner SA, Sokoli A, Felder KM, et al. (2012) The surface-localised α-enolase of Mycoplasma suis is an adhesion protein. Vet Microbiol 156: 88–95. doi: 10.1016/j.vetmic.2011.10.010
|
| [85] |
Matta SK, Agarwal S, Bhatnagar R (2010) Surface localized and extracellular Glyceraldehyde-3-phosphate dehydrogenase of Bacillus anthracis is a plasminogen binding protein. BBA-Proteins Proteom 1804: 2111–2120. doi: 10.1016/j.bbapap.2010.08.004
|
| [86] |
Seifert KN, McArthur WP, Bleiweis AS, et al. (2003) Characterization of group B streptococcal glyceraldehyde-3-phosphate dehydrogenase: surface localization, enzymatic activity, and protein-protein interactions. Can J Microbiol 49: 350–356. doi: 10.1139/w03-042
|
| [87] |
Bergmann S, Rohde M, Hammerschmidt S (2004) Glyceraldehyde-3-phosphate dehydrogenase of Streptococcus pneumoniae is a surface-displayed plasminogen-binding protein. Infect Immun 72: 2416–2419. doi: 10.1128/IAI.72.4.2416-2419.2004
|
| [88] |
Winram SB, Lottenberg R (1996) The plasmin-binding protein Plr of group A streptococci is identified as glyceraldehyde-3-phosphate dehydrogenase. Microbiology 142: 2311–2320. doi: 10.1099/13500872-142-8-2311
|
| [89] |
Jobin MC, Brassard J, Quessy S, et al. (2004) Acquisition of host plasmin activity by the Swine pathogen Streptococcus suis serotype 2. Infect Immun 72: 606–610. doi: 10.1128/IAI.72.1.606-610.2004
|
| [90] |
Kainulainen V, Loimaranta V, Pekkala A, et al. (2012) Glutamine synthetase and glucose-6-phosphate isomerase are adhesive moonlighting proteins of Lactobacillus crispatus released by epithelial cathelicidin LL-37. J Bacteriol 194: 2509–2519. doi: 10.1128/JB.06704-11
|
| [91] |
Xolalpa W, Vallecillo AJ, Lara M, et al. (2007) Identification of novel bacterial plasminogen-binding proteins in the human pathogen Mycobacterium tuberculosis. Proteomics 7: 3332–3341. doi: 10.1002/pmic.200600876
|
| [92] | Hickey TB, Thorson LM, Speert DP, et al. (2009) Mycobacterium tuberculosis Cpn60.2 and DnaK are located on the bacterial surface, where Cpn60.2 facilitates efficient bacterial association with macrophages. Infect Immun 77: 3389–3401. |
| [93] |
Jarocki VM, Santos J, Tacchi JL, et al. (2015) MHJ_0461 is a multifunctional leucine aminopeptidase on the surface of Mycoplasma hyopneumoniae. Open Biol 5: 140175. doi: 10.1098/rsob.140175
|
| [94] | Hussain M, Peters G, Chhatwal GS, et al. (1999) A lithium chloride-extracted, broad-spectrum-adhesive 42-kilodalton protein of Staphylococcus epidermidis is ornithine carbamoyltransferase. Infect Immun 67: 6688–6690. |
| [95] | Lechardeur D, Fernandez A, Robert B, et al. (2011) The 2-Cys peroxiredoxin alkyl hydroperoxide reductase c binds heme and participates in its intracellular availability in Streptococcus agalactiae. J Biol Chem 285: 16032–16041. |
| [96] |
Boone TJ, Burnham CA, Tyrrell GJ (2011) Binding of group B streptococcal phosphoglycerate kinase to plasminogen and actin. Microb Pathogenesis 51: 255–261. doi: 10.1016/j.micpath.2011.06.005
|
| [97] |
Fulde M, Bernardo-Garcia N, Rohde M, et al. (2014) Pneumococcal phosphoglycerate kinase interacts with plasminogen and its tissue activator. Thromb Haemostasis 111: 401–416. doi: 10.1160/TH13-05-0421
|
| [98] |
Reddy VM, Suleman FG (2004) Mycobacterium avium superoxide dismutase binds to epithelial cell aldolase, glyceraldehyde-3-phosphate dehydrogenase and cyclophilin A. Microb Pathogenesis 36: 67–74. doi: 10.1016/j.micpath.2003.09.005
|
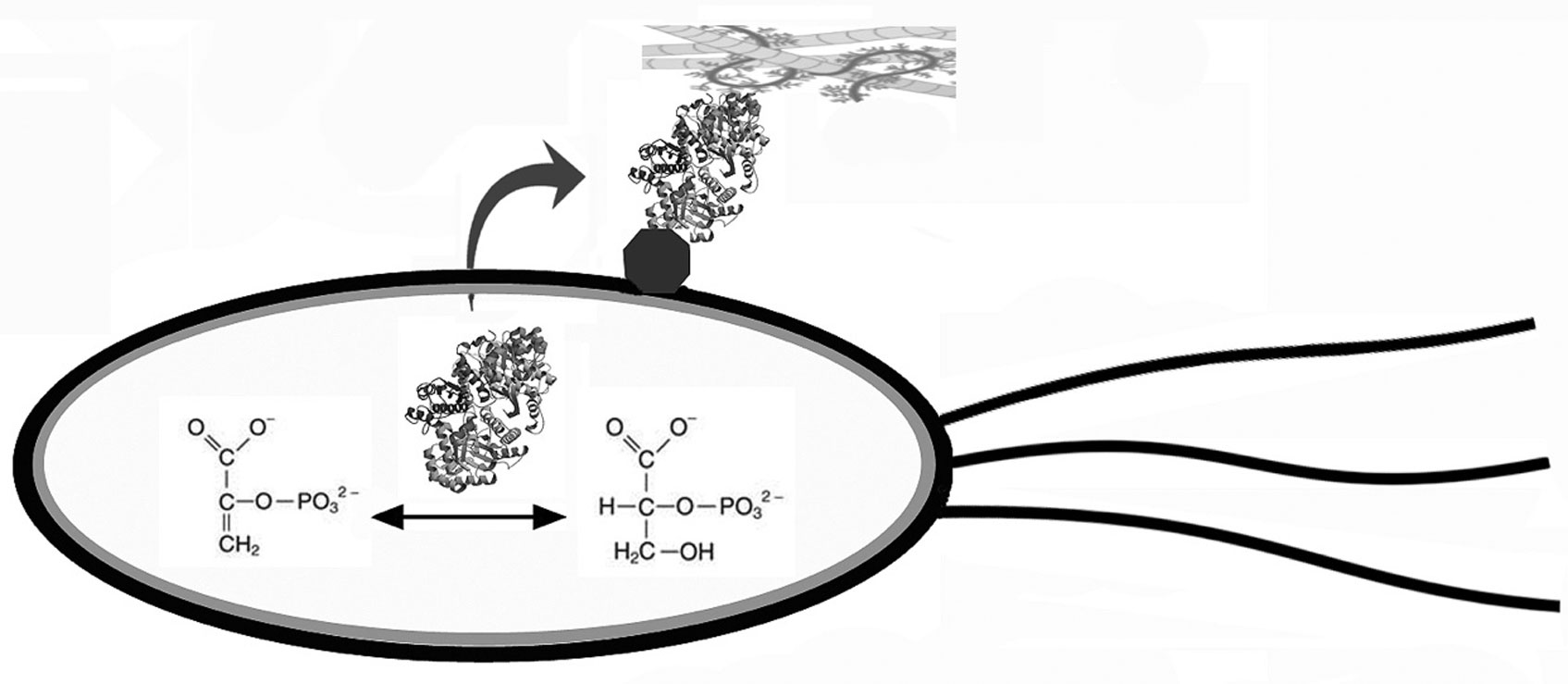
Figures(1) / Tables(1)

Constance Jeffery. Intracellular proteins moonlighting as bacterial adhesion factors[J]. AIMS Microbiology, 2018, 4(2): 362-376. doi: 10.3934/microbiol.2018.2.362







 DownLoad:
DownLoad: