Citation: Luciana De Vero, Tommaso Bonciani, Alexandra Verspohl, Francesco Mezzetti, Paolo Giudici. High-glutathione producing yeasts obtained by genetic improvement strategies: a focus on adaptive evolution approaches for novel wine strains[J]. AIMS Microbiology, 2017, 3(2): 155-170. doi: 10.3934/microbiol.2017.2.155

| [1] |
Pretorius IS (2000) Tailoring wine yeast for the new millennium: novel approaches to the ancient art of winemaking. Yeast 16: 675–729. doi: 10.1002/1097-0061(20000615)16:8<675::AID-YEA585>3.0.CO;2-B

|
| [2] |
Giudici P, Solieri L, Pulvirenti A, et al. (2005) Strategies and perspectives for genetic improvement of wine yeasts. Appl Microbiol Biotechnol 66: 622–628. doi: 10.1007/s00253-004-1784-2
|
| [3] | Giudici P, Zambonelli C (1992) Criteri di selezione di lieviti per enologia. Vignevini 9: 29–34 [In Italian]. |
| [4] | Rainieri S, Zambonelli C, Tini V, et al. (1998) The enological traits of thermotolerant Saccharomyces Strains. Am J Enol Vitic 49: 319–324. |
| [5] | Anderson ME (1998) Glutathione: An overview of biosynthesis and modulation. Chem Biol Interact 111: 1–14. |
| [6] | Ganguly D, Srikanth CV, Kumar C, et al. (2003) Why is glutathione (a tripeptide) synthesized by specific enzymes while TSH releasing hormone (TRH or thyroliberin), also a tripeptide, is produced as part of a prohormone protein? IUBMB Life 56: 361–363. |
| [7] |
Meister A, Anderson ME (1983) Glutathione. Annu Rev Biochem 52: 711–760. doi: 10.1146/annurev.bi.52.070183.003431
|
| [8] |
Kritzinger EC, Bauer FF, du Toit WJ (2013) Role of glutathione in winemaking: a review. J Agric Food Chem 61: 269–277. doi: 10.1021/jf303665z
|
| [9] |
Oliveira CM, Ferreira ACS, De Freitas V, et al. (2011) Oxidation mechanisms occurring in wines. Food Res Int 44: 1115–1126. doi: 10.1016/j.foodres.2011.03.050
|
| [10] |
Sonni F, Clark AC, Prenzler PD, et al. (2011) Antioxidant action of glutathione and the ascorbic acid/glutathione pair in a model white wine. J Agric Food Chem 59: 3940–3949. doi: 10.1021/jf104575w
|
| [11] |
Sonni F, Moore EG, Clark AC, et al. (2011) Impact of glutathione on the formation of methylmethine-and carboxymethine-bridged (+)-catechin dimers in a model wine system. J Agric Food Chem 59: 7410–7418. doi: 10.1021/jf200968x
|
| [12] | Dubourdieu D, Lavigne V (2004) The role of glutathione on the aromatic evolution of dry white wine. Vinidea.net 2: 1–9. |
| [13] | Roussis I, Papadopoulou D, Sakarellos-Daitsiotis M (2009) Protective effect of thiols on wine aroma volatiles. Open Food Sci J 3: 98–102. |
| [14] |
Tirelli A, Fracassetti D, De Noni I (2010) Determination of reduced cysteine in oenological cell wall fractions of Saccharomyces cerevisiae. J Agric Food Chem 58: 4565–4570. doi: 10.1021/jf904047u
|
| [15] | Park SK, Boulton R, Noble A (2000) Formation of hydrogen sulfide and glutathione during fermentation of white grape musts. Am J Enol Vitic 51: 91–97. |
| [16] |
Kritzinger EC, Bauer FF, Du Toit WJ (2013) Influence of yeast strain, extended lees contact and nitrogen supplementation on glutathione concentration in wine. Aust J Grape Wine Res 19: 161–170. doi: 10.1111/ajgw.12025
|
| [17] | OIV, RESOLUTION OIV-OENO 445-2015, 2015. Available from: http://www.oiv.int/en/technical-standards-and-documents/resolutions-oiv/oenology-resolutions. |
| [18] | OIV, RESOLUTION OIV-OENO 446-2015, 2015. Available from: http://www.oiv.int/en/technical-standards-and-documents/resolutions-oiv/oenology-resolutions. |
| [19] |
Wegmann-Herr P, Ullrich S, Schmarr HG, et al. (2016) Use of glutathione during white wine production- impact on S-off-flavors and sensory production. BIO Web Conf 7: 2031. doi: 10.1051/bioconf/20160702031
|
| [20] |
Pozo-Bayón MÁ, Andújar-Ortiz I, Moreno-Arribas MV (2009) Scientific evidences beyond the application of inactive dry yeast preparations in winemaking. Food Res Int 42: 754–761. doi: 10.1016/j.foodres.2009.03.004
|
| [21] | Andujar-Ortiz I, Pozo-Bayón MÁ, Moreno-Arribas MV, et al. (2012) Reversed-phase high-performance liquid chromatography–fluorescence detection for the analysis of glutathione and its precursor γ-glutamyl cysteine in wines and model wines supplemented with oenological inactive dry yeast preparations. Food Anal Methods 5: 154–161. |
| [22] | Kritzinger E, Stander M, Du Toit W (2013) Assessment of glutathione levels in model solution and grape ferments supplemented with glutathione-enriched inactive dry yeast preparations using a novel UPLC-MS/MS method. Food Addit Contam 30: 37–41. |
| [23] |
Elskens M, Jaspers C, Penninckx M (1991) Glutathione as an endogenous sulphur source in the yeast Saccharomyces cerevisiae. J Gen Microbiol 137: 637–644. doi: 10.1099/00221287-137-3-637
|
| [24] | Thomas D, Surdin-Kerjan Y (1997) Metabolism of sulfur amino acids in Saccharomyces cerevisiae. Microbiol Mol Biol Rev 61: 503–532. |
| [25] | Mendoza-Cózatl D, Loza-Tavera H, Hernández-Navarro A, et al. (2005) Sulfur assimilation and glutathione metabolism under cadmium stress in yeast, protists and plants. FEMS Microbiol Rev 29: 653–671. |
| [26] | Penninckx MJ (2002) An overview on glutathione in Saccharomyces versus non-conventional yeasts. FEMS Yeast Res 2: 295–305. |
| [27] |
Suzuki T, Yokoyama A, Tsuji T, et al. (2011) Identification and characterization of genes involved in glutathione production in yeast. J Biosci Bioeng 112: 107–113. doi: 10.1016/j.jbiosc.2011.04.007
|
| [28] |
Bourbouloux A, Shahi P, Chakladar A, et al. (2000) Hgt1p, A high affinity glutathione transporter from the yast Saccharomyces cerevisiae. J Biol Chem 275: 13259–13265. doi: 10.1074/jbc.275.18.13259
|
| [29] |
Dhaoui M, Auchère F, Blaiseau PL, et al. (2011) Gex1 is a yeast glutathione exchanger that interferes with pH and redox homeostasis. Mol Biol Cell 22: 2054–2067. doi: 10.1091/mbc.E10-11-0906
|
| [30] |
Mehdi K, Penninckx MJ (1997) An important role for glutathione and Υ-glutamyltranspeptidase in the supply of growth requirements during nitrogen starvation of the yeast Saccharomyces cerevisiae. Microbiology 143: 1885–1889. doi: 10.1099/00221287-143-6-1885
|
| [31] |
Grant CM (2001) Role of the glutathione/glutaredoxin and thioredoxin systems in yeast growth and response to stress conditions. Mol Microbiol 39: 533–541. doi: 10.1046/j.1365-2958.2001.02283.x
|
| [32] |
Izawa S, Inoue Y, Kimura A (1995) Oxidative stress-response in yeast: effect of glutathione on adaptation to hydrogen-peroxide stress in Saccharomyces cerevisiae. FEBS Lett 368: 73–76. doi: 10.1016/0014-5793(95)00603-7
|
| [33] |
Penninckx M (2000) A short review on the role of glutathione in the response of yeasts to nutritional, environmental, and oxidative stresses. Enzyme Microb Technol 26: 737–742. doi: 10.1016/S0141-0229(00)00165-4
|
| [34] |
Mortimer RK (2000) Evolution and variation of the yeast (Saccharomyces) genome. Genome Res 10: 403–409. doi: 10.1101/gr.10.4.403
|
| [35] |
Sipiczki M (2011) Diversity, variability and fast adaptive evolution of the wine yeast (Saccharomyces cerevisiae) genome-a review. Ann Microbiol 61: 85–93. doi: 10.1007/s13213-010-0086-4
|
| [36] | Romano P, Soli MG, Suzzi G (1983) Procedure for mutagenizing spores of Saccharomyces cerevisiae. J Bacteriol 156: 907–908. |
| [37] |
Rous CV, Snow R, Kunkee RE (1983) Reduction of higher alcohols by fermentation with a leucine-auxotrophic mutant of wine yeast. J Inst Brew 89: 274–278. doi: 10.1002/j.2050-0416.1983.tb04185.x
|
| [38] | Giudici P, Zinnato A (1983) Influenza dell'uso di mutanti nutrizionali sulla produzione di alcooli superiori. Vignevini 10: 63–65 [In Italian]. |
| [39] | Ikeno Y, Tanno K, Omori I, et al. (1977) Glutathione. Japanese Patent Application 52(087): 296. |
| [40] | Kono G, Harada M, Sugisaki K, et al. (1977) High glutathione-containing yeast. Japanese Patent Application 52(125): 687. |
| [41] | Hamada S, Tanaka H, Sakato K (1986) Process for producing glutathione. United States Patent Application 4(582): 801. |
| [42] |
Li Y, Wei G, Chen J (2004) Glutathione: a review on biotechnological production. Appl Microbiol Biotechnol 66: 233–242. doi: 10.1007/s00253-004-1751-y
|
| [43] |
Nisamedtinov I, Kevvai K, Orumets K, et al. (2011) Metabolic changes underlying the higher accumulation of glutathione in Saccharomyces cerevisiae mutants. Appl Microbiol Biotechnol 89: 1029–1037. doi: 10.1007/s00253-010-2946-z
|
| [44] |
Steensels J, Snoek T, Meersman E, et al. (2014) Improving industrial yeast strains: exploiting natural and artificial diversity. FEMS Microbiol Rev 38: 947–995. doi: 10.1111/1574-6976.12073
|
| [45] |
Zambonelli C, Passarelli P, Rainieri S, et al. (1997) Technological properties and temperature response of interspecific Saccharomyces hybrids. J Sci Food Agric 74: 7–12. doi: 10.1002/(SICI)1097-0010(199705)74:1<7::AID-JSFA753>3.0.CO;2-X
|
| [46] |
Rainieri S, Zambonelli C, Giudici P, et al. (1998) Characterisation of thermotolerant Saccharomyces cerevisiae hybrids. Biotechnol Lett 20: 543–547. doi: 10.1023/A:1005389309527
|
| [47] |
Bellon JR, Eglinton JM, Siebert TE, et al. (2011) Newly generated interspecific wine yeast hybrids introduce flavour and aroma diversity to wines. Appl Microbiol Biotechnol 91: 603–612. doi: 10.1007/s00253-011-3294-3
|
| [48] |
Bonciani T, Solieri L, De Vero L, et al. (2016) Improved wine yeasts by direct mating and selection under stressful fermentative conditions. Eur Food Res Technol 242: 899–910. doi: 10.1007/s00217-015-2596-6
|
| [49] | Freese EB, Chu MI, Freese E (1982) Initiation of yeast sporulation by partial carbon, nitrogen, or phosphate deprivation. J Bacteriol 149: 840–851. |
| [50] |
Solieri L, Verspohl A, Bonciani T, et al. (2015) Fast method for identifying inter- and intra-species Saccharomyces hybrids in extensive genetic improvement programs based on yeast breeding. J Appl Microbiol 119: 149–161. doi: 10.1111/jam.12827
|
| [51] |
Verspohl A, Solieri L, Giudici P (2017) Exploration of genetic and phenotypic diversity within Saccharomyces uvarum for driving strain improvement in winemaking. Appl Microbiol Biotechnol 101: 2507–2521. doi: 10.1007/s00253-016-8008-4
|
| [52] |
Krivoruchko A, Siewers V, Nielsen J (2011) Opportunities for yeast metabolic engineering: lessons from synthetic biology. Biotechnol J 6: 262–276. doi: 10.1002/biot.201000308
|
| [53] | Goffeau A, Barrell BG, Bussey H, et al. (1996) Life with 6000 Genes. Science 274 (5287): 546–567. |
| [54] |
Pérez-Torrado R, Querol A, Guillamón JM (2015) Genetic improvement of non-GMO wine yeasts: Strategies, advantages and safety. Trends Food Sci Technol 45: 1–11. doi: 10.1016/j.tifs.2015.05.002
|
| [55] | Hosiner D, Gerber S, Lichtenberg-Fraté H, et al. (2014) Impact of acute metal stress in Saccharomyces cerevisiae. PLoS One 9: 1–14. |
| [56] |
Collinson LP, Dawes IW (1995) Isolation, characterization and overexpression of the yeast gene, GLR1, encoding glutathione reductase. Gene 156: 123–127. doi: 10.1016/0378-1119(95)00026-3
|
| [57] |
Grant CM, MacIver FH, Dawes IW (1997) Glutathione synthetase is dispensable for growth under both normal and oxidative stress conditions in the yeast Saccharomyces cerevisiae due to an accumulation of the dipeptide y-glutamylcysteine. Mol Biol Cell 8: 1699–1707. doi: 10.1091/mbc.8.9.1699
|
| [58] |
Hara KY, Kiriyama K, Inagaki A, et al. (2012) Improvement of glutathione production by metabolic engineering the sulfate assimilation pathway of Saccharomyces cerevisiae. Appl Microbiol Biotechnol 94: 1313–1319. doi: 10.1007/s00253-011-3841-y
|
| [59] |
Ask M, Mapelli V, Höck H, et al. (2013) Engineering glutathione biosynthesis of Saccharomyces cerevisiae increases robustness to inhibitors in pretreated lignocellulosic materials. Microb Cell Fact 12: 87. doi: 10.1186/1475-2859-12-87
|
| [60] |
Qiu Z, Deng Z, Tan H, et al. (2015) Engineering the robustness of Saccharomyces cerevisiae by introducing bifunctional glutathione synthase gene. J Ind Microbiol Biotechnol 42: 537–542. doi: 10.1007/s10295-014-1573-6
|
| [61] | Sauer U (2001) Evolutionary engineering of industrially important microbial phenotypes, In: Metabolic Engineering, Springer Berlin Heidelberg, 129–169. |
| [62] | McBryde C, Gardner JM, De Barros Lopes M, et al. (2006) Generation of novel wine yeast strains by adaptive evolution. Am J Enol Vitic 57: 423–430. |
| [63] |
De Vero L, Solieri L, Giudici P (2011) Evolution-based strategy to generate non-genetically modified organisms Saccharomyces cerevisiae strains impaired in sulfate assimilation pathway. Lett Appl Microbiol 53: 572–575. doi: 10.1111/j.1472-765X.2011.03140.x
|
| [64] |
Winkler JD, Kao KC (2014) Recent advances in the evolutionary engineering of industrial biocatalysts. Genomics 104: 406–411. doi: 10.1016/j.ygeno.2014.09.006
|
| [65] |
Brown SW, Oliver SG (1982) Isolation of ethanol-tolerant mutants of yeast by continuous selection. Eur J Appl Microbiol Biotechnol 16: 119–122. doi: 10.1007/BF00500738
|
| [66] |
Cakar ZP, Seker UOS, Tamerler C, et al. (2005) Evolutionary engineering of multiple-stress resistant Saccharomyces cerevisiae. FEMS Yeast Res. 5: 569–578. doi: 10.1016/j.femsyr.2004.10.010
|
| [67] |
Cadière A, Ortiz-Julien A, Camarasa C, et al. (2011) Evolutionary engineered Saccharomyces cerevisiae wine yeast strains with increased in vivo flux through the pentose phosphate pathway. Metab Eng 13: 263–271. doi: 10.1016/j.ymben.2011.01.008
|
| [68] |
Kutyna DR, Varela C, Stanley GA, et al. (2012) Adaptive evolution of Saccharomyces cerevisiae to generate strains with enhanced glycerol production. Appl Microbiol Biotechnol 93: 1175–1184. doi: 10.1007/s00253-011-3622-7
|
| [69] |
Mezzetti F, De Vero L, Giudici P (2014) Evolved Saccharomyces cerevisiae wine strains with enhanced glutathione production obtained by an evolution-based strategy. FEMS Yeast Res 14: 977–987. doi: 10.1111/1567-1364.12186
|
| [70] |
Cakar ZP, Turanli-Yildiz B, Alkim C, et al. (2012) Evolutionary engineering of Saccharomyces cerevisiae for improved industrially important properties. FEMS Yeast Res 12: 171–182. doi: 10.1111/j.1567-1364.2011.00775.x
|
| [71] | Cherest H, Davidian J, Benes V, et al. (1997) Molecular characterization of two high affinity sulfate transporters in Saccharomyces cerevisiae. Genet Soc Am 145: 627–635. |
| [72] | Tamás MJ, Labarre J, Toledano MB, et al. (2005) Mechanisms of toxic metal tolerance in yeast, In: Tamás M.J., Martinoia E., Molecular biology of metal homeostasis and detoxification: from microbes to man, Heidelberg: Springer Verlag, 395–454. |
| [73] |
Wysocki R, Tamás MJ (2010) How Saccharomyces cerevisiae copes with toxic metals and metalloids. FEMS Microbiol Rev 34: 925–951. doi: 10.1111/j.1574-6976.2010.00217.x
|
| [74] |
Zechmann B, Liou LC, Koffler BE, et al. (2011) Subcellular distribution of glutathione and its dynamic changes under oxidative stress in the yeast Saccharomyces cerevisiae. FEMS Yeast Res 11: 631–642. doi: 10.1111/j.1567-1364.2011.00753.x
|
| [75] | Ortiz DF, Kreppel L, Speiser DM, et al. (1992) Heavy metal tolerance in the fission yeast requires an ATP-binding cassette-type vacuolar membrane transporter. EMBO J 11: 3491–3499. |
| [76] |
Duncan WS, Jamieson DJ (1996) Glutathione is an important antioxidant molecule in the yeast Saccharomyces cerevisiae. FEMS Microbiol Lett 141: 207–212. doi: 10.1111/j.1574-6968.1996.tb08386.x
|
| [77] |
Adamis PDB, Panek AD, Eleutherio ECA (2007) Vacuolar compartmentation of the cadmium-glutathione complex protects Saccharomyces cerevisiae from mutagenesis. Toxicol Lett 173: 1–7. doi: 10.1016/j.toxlet.2007.06.002
|
| [78] |
Patzschke A, Steiger MG, Holz C, et al. (2015) Enhanced glutathione production by evolutionary engineering of Saccharomyces cerevisiae strains. Biotechnol J 10: 1719–1726. doi: 10.1002/biot.201400809
|
| [79] |
Bailey JE, Sburlati A, Hatzimanikatis V, et al. (2002) Inverse metabolic engineering: a strategy for directed genetic engineering of useful phenotypes. Biotechnol Bioeng 79: 568–579. doi: 10.1002/bit.10441
|
| [80] |
Rossouw D, Naes T, Bauer FF (2008) Linking gene regulation and the exo-metabolome: a comparative transcriptomics approach to identify genes that impact on the production of volatile aroma compounds in yeast. BMC Genomics 9: 530. doi: 10.1186/1471-2164-9-530
|
| [81] | Liti G, Louis EJ (2012) Advances in quantitative trait analysis in yeast. PLoS Genet 8: 1–7. |
| [82] |
Fleet GH (2003) Yeast interactions and wine flavour. Int J Food Microbiol 86: 11–22. doi: 10.1016/S0168-1605(03)00245-9
|
| [83] |
Pizarro F, Vargas FA, Agosin E (2007) A systems biology perspective of wine fermentations. Yeast 24: 977–991. doi: 10.1002/yea.1545
|
| [84] |
Siezen RJ (2008) Wine genomics. Microb Biotechnol 1: 97–103. doi: 10.1111/j.1751-7915.2008.00030.x
|
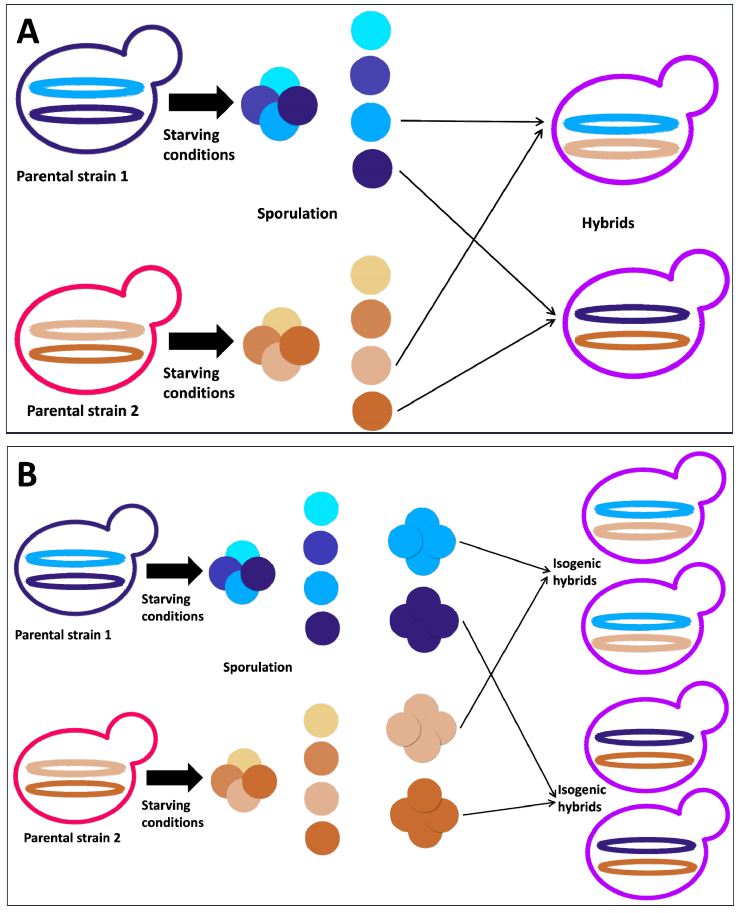
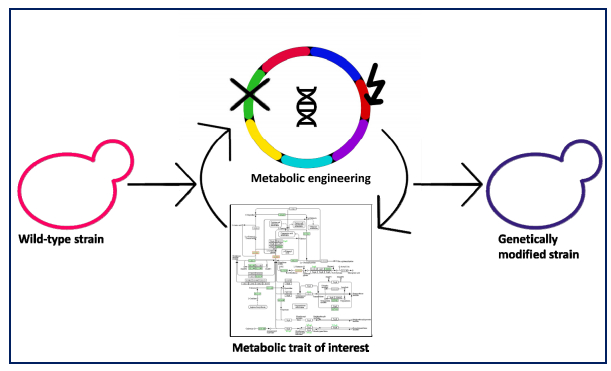
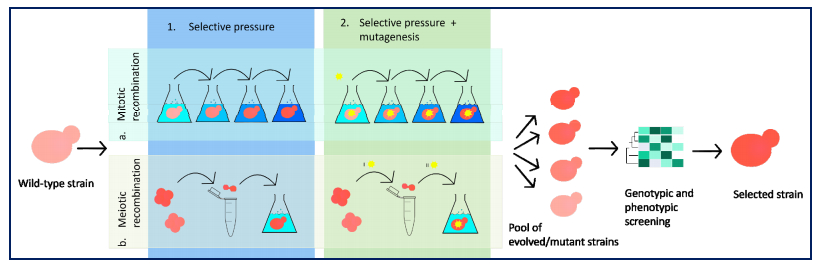
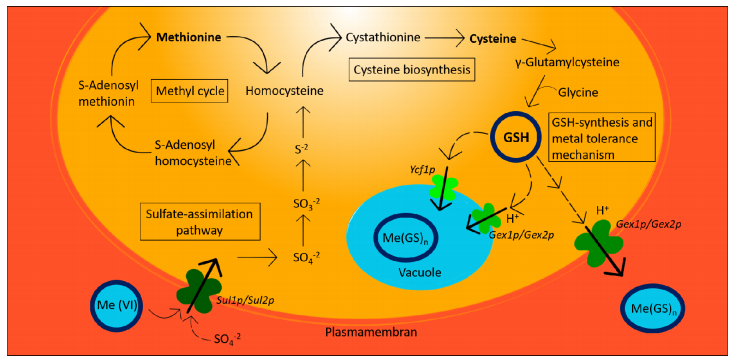
Figures(4)

Luciana De Vero, Tommaso Bonciani, Alexandra Verspohl, Francesco Mezzetti, Paolo Giudici. High-glutathione producing yeasts obtained by genetic improvement strategies: a focus on adaptive evolution approaches for novel wine strains[J]. AIMS Microbiology, 2017, 3(2): 155-170. doi: 10.3934/microbiol.2017.2.155







 DownLoad:
DownLoad: