Citation: Hsiu-Chuan Wei. Mathematical modeling of tumor growth: the MCF-7 breast cancer cell line[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6512-6535. doi: 10.3934/mbe.2019325

| [1] | D. R. Jutagir, B. B. Blomberg, C. S. Carver, et al., Social well-being is associated with less pro-inflammatory and pro-metastatic leukocyte gene expression in women after surgery for breast cancer, Breast Cancer Res. Treat., 165 (2017), 169–180. |
| [2] | S. Katkuri and M. Gorantla, Awareness about breast cancer among women aged 15 years and above in urban slums: a cross sectional study, Int. J. Community Med. Public Health, 5 (2018), 929–932. |
| [3] | A. Pawlik, M. Slomi´ nska-Wojewódzka and A. Herman-Antosiewicz, Sensitization of estrogen receptor-positive breast cancer cell lines to 4-hydroxytamoxifen by isothiocyanates present in cruciferous plants, Eur. J. Nutr., 55 (2016), 1165–1180. |
| [4] | D. L. Holliday and V. Speirs, Choosing the right cell line for breast cancer research, Breast Cancer Res., 13 (2011), 215–215. |
| [5] | R. L. Sutherland, R. E. Hall and I. W. Taylor, Cell proliferation kinetics of MCF-7 human mammary carcinoma cells in culture and effects of tamoxifen on exponentially growing and plateau-phase cells, Cancer Res., 43 (1983), 3998–4006. |
| [6] | B. S. Katzenellenbogen, K. L. Kendra, M. J. Norman, et al., Proliferation, hormonal responsiveness, and estrogen receptor content of MCF-7 human breast cancer cells grown in the short-term and long-term absence of estrogens, Cancer Res., 47(1987), 4355–4360. |
| [7] | A. Maton, Human biology and health, 1st edition, Prentice Hall, New Jersey, 1997. |
| [8] | L. V. Rao, B. A. Ekberg, D. Connor, et al., Evaluation of a new point of care automated complete blood count (CBC) analyzer in various clinical settings, Clin. Chim. Acta., 389 (2008), 120–125. |
| [9] | S. Bernard, E. Abdelsamad, P. Johnson, et al., Pediatric leukemia: diagnosis to treatment a review, J. Cancer Clin. Trials, 2(2017), 131. |
| [10] | A. Shankar, J. J. Wang, E. Rochtchina, et al., Association between circulating white blood cell count and cancer mortality: a population-based cohort study, Arch. Intern. Med., 166 (2006), 188–194. |
| [11] | K. Kim, J. Lee, N. J. Heo, et al., Differential white blood cell count and all-cause mortality in the Korean elderly, Exp. Gerontol., 48 (2013), 103–108. |
| [12] | C. Ruggiero, E. J. Metter, A. Cherubini, et al., White blood cell count and mortality in the Baltimore Longitudinal Study of Aging, J. Am. Coll. Cardiol., 49 (2007), 1841–1850. |
| [13] | D. S. Bell and J. H. O'Keefe, White cell count, mortality, and metabolic syndrome in the Baltimore longitudinal study of aging, J. Am. Coll. Cardiol., 50(2007), 1810. |
| [14] | G. D. Friedman and B. H. Fireman, The leukocyte count and cancer mortality, Am. J. Epidemiol., 133 (1991), 376–380. |
| [15] | M. H. Andersen, D. Schrama, P. thor Straten, et al., Cytotoxic T cells, J. Invest. Dermatol., 126 (2006), 32–41. |
| [16] | J. Folkman and R. Kalluri, Cancer without disease, Nature, 427 (2004), 787. |
| [17] | T. Fehm, V. Mueller, R. Marches, et al., Tumor cell dormancy: implications for the biology and treatment of breast cancer, Apmis, 116 (2008), 742–753. |
| [18] | N. Almog, Molecular mechanisms underlying tumor dormancy, Cancer Lett., 294 (2010), 139–146. |
| [19] | A. Friedman, Cancer as multifaceted disease, Math. Model. Nat. Pheno., 7(2012), 3–28. |
| [20] | R. Eftimie, J. L. Bramson and D. J. Earn, Interactions between the immune system and cancer: a brief review of non-spatial mathematical models, Bull. Math. Biol., 73 (2011), 2–32. |
| [21] | S. Banerjee and R. R. Sarkar, Delay-induced model for tumor–immune interaction and control of malignant tumor growth, Biosystems, 91 (2008), 268–288. |
| [22] | H. Moore and N. K. Li, A mathematical model for chronic myelogenous leukemia (CML) and T cell interaction, J. Theor. Biol., 227 (2004), 513–523. |
| [23] | L. Anderson, S. Jang and J. Yu, Qualitative behavior of systems of tumor-CD4+-cytokine interactions with treatments, Math. Method. Appl. Sci., 38 (2015), 4330–4344. |
| [24] | A. d'Onofrio, Metamodeling tumor–immune system interaction, tumor evasion and immunotherapy, Math. Comput. Model., 47 (2008), 614–637. |
| [25] | A. Khar, Mechanisms involved in natural killer cell mediated target cell death leading to spontaneous tumour regression, J. Biosci., 22 (1997), 23–31. |
| [26] | T. Boon and P. van der Bruggen, Human tumor antigens recognized by T lymphocytes, J. Exp. Med., 183 (1996), 725–729. |
| [27] | D. Kirschner and J. C. Panetta, Modeling immunotherapy of the tumor–immune interaction, J. Math. Biol., 37 (1998), 235–252. |
| [28] | L. G. de Pillis, W. Gu and A. E. Radunskaya, Mixed immunotherapy and chemotherapy of tumors: modeling, applications and biological interpretations, J. Theor. Biol., 238 (2006), 841–862. |
| [29] | H. P. de Vladar and J. A. González, Dynamic response of cancer under the influence of immunological activity and therapy, J. Theor. Biol., 227 (2004), 335–348. |
| [30] | U. Fory´ s, J. Waniewski and P. Zhivkov, Anti-tumor immunity and tumor anti-immunity in a mathematical model of tumor immunotherapy, J. Biol. Syst., 14 (2006), 13–30. |
| [31] | R. W. De Boer, J. M. Karemaker and J. Strackee, Relationships between short-term blood-pressure fluctuations and heart-rate variability in resting subjects I: a spectral analysis approach, Med. Biol. Eng. Comput., 23 (1985), 352–358. |
| [32] | A. Cappuccio, M. Elishmereni and Z. Agur, Cancer immunotherapy by interleukin-21: potential treatment strategies evaluated in a mathematical model, Cancer Res., 66 (2006), 7293–7300. |
| [33] | N. Kronik, Y. Kogan, V. Vainstein, et al., Improving alloreactive CTL immunotherapy for malignant gliomas using a simulation model of their interactive dynamics, Cancer Immunol. Immunother., 57 (2008), 425–439. |
| [34] | A. M. Jarrett, J. M. Bloom, W. Godfrey, et al., Mathematical modelling of trastuzumab-induced immune response in an in vivo murine model of HER2+ breast cancer, Math. Med. Biol., (2018), dqy014. |
| [35] | K. Annan, M. Nagel and H. A. Brock, A mathematical model of breast cancer and mediated immune system interactions, J. Math. Syst. Sci., 2(2012), 430–446. |
| [36] | R. Roe-Dale, D. Isaacson and M. Kupferschmid, A mathematical model of breast cancer treatment with CMF and doxorubicin, Bull. Math. Biol., 73 (2011), 585–608. |
| [37] | B. Ribba, N. H. Holford, P. Magni, et al., A review of mixed-effects models of tumor growth and effects of anticancer drug treatment used in population analysis, CPT Pharmacometrics Syst. Pharmacol., 3(2014), 1–10. |
| [38] | R. Bhat and C. Watzl, Serial killing of tumor cells by human natural killer cells–enhancement by therapeutic antibodies, PloS One, 2 (2007), e326. |
| [39] | T. Sutlu and E. Alici, Natural killer cell-based immunotherapy in cancer: current insights and future prospects, J. Intern. Med., 266 (2009), 154–181. |
| [40] | T. R. Stravitz, T. Lisman, V. A. Luketic, et al., Minimal effects of acute liver injury/acute liver failure on hemostasis as assessed by thromboelastography, J. Hepatol., 56 (2012), 129–136. |
| [41] | Y. Zhang, D. L. Wallace, C. M. De Lara, et al., In vivo kinetics of human natural killer cells: the effects of ageing and acute and chronic viral infection, Immunotherapy, 121(2007), 258–265. |
| [42] | P. Wilding, L. J. Kricka, J. Cheng, et al., Integrated cell isolation and polymerase chain reaction analysis using silicon microfilter chambers, Anal. Biochem., 257(1998), 95–100. |
| [43] | V. Pascal, N. Schleinitz, C. Brunet, et al., Comparative analysis of NK cell subset distribution in normal and lymphoproliferative disease of granular lymphocyte conditions, Eur. J. Immunol., 34(2004), 2930–2940. |
| [44] | L. de Pillis, T. Caldwell, E. Sarapata, et al., Mathematical modeling of the regulatory T cell effects on renal cell carcinoma treatment, Discrete Continuous Dyn. Syst. Ser. B, 18(2013), 915–943. |
| [45] | T. D. To, A. T. T. Truong, A. T. Nguyen, et al., Filtration of circulating tumour cells MCF-7 in whole blood using non-modified and modified silicon nitride microsieves, Int. J. Nanotechnol., 15(2018), 39–52. |
| [46] | C. Chen, Y. Chen, D. Yao, et al., Centrifugalfilter device for detection of rare cells with immuno-binding, IEEE T. Nanobiosci., 14(2015), 864–869. |
| [47] | P. Dua, V. Dua and E. N. Pistikopoulos, Optimal delivery of chemotherapeutic agents in cancer, Comput. Chem. Eng., 32(2008), 99–107. |
| [48] | A. G. López, J. M. Seoane and M. A. Sanjuán, A validated mathematical model of tumor growth including tumor–host interaction, cell-mediated immune response and chemotherap, Bull. Math. Biol., 76(2014), 2884–2906. |
| [49] | K. Liao, X. Bai and A. Friedman, The role of CD200–CD200R in tumor immune evasion, J. Theor. Biol., 328(2013), 65–76. |
| [50] | M. C. Martins, A. M. A. Rocha, M. F. P. Costa, et al., Comparing immune-tumor growth models with drug therapy using optimal control, AIP Conf. Proc., 1738(2016), 300005. |
| [51] | M. Fernandez, M. Zhou and L. Soto-Ortiz, A computational assessment of the robustness of cancer treatments with respect to immune response strength, tumor size and resistance, Int. J. Tumor Ther., 7(2018), 1–26. |
| [52] | D. F. Tough and J. Sprent, Life span of naive and memory T cells, Stem Cells, 13(1995), 242–249. |
| [53] | C. M. Rollings, L. V. Sinclair, H. J. M. Brady, et al., Interleukin-2 shapes the cytotoxic T cell proteome and immune environment–sensing programs, Sci. Signal., 11(2018), eaap8112. |
| [54] | E. M. Janssen, E. E. Lemmens, T. Wolfe, et al., CD4+ T cells are required for secondary expansion and memory in CD8+ T lymphocytes, Nature, 421(2003), 852. |
| [55] | I. Gruber, N. Landenberger, A. Staebler, et al., Relationship between circulating tumor cells and peripheral T-cells in patients with primary breast cancer, Anticancer Res., 33(2013), 2233–2238. |
| [56] | D. Homann, L. Teyton and M. B. Oldstone, Differential regulation of antiviral T-cell immunity results in stable CD8+ but declining CD4+ T-cell memory, Nat. Med., 7(2001), 913–919. |
| [57] | R. J. De Boer, D. Homann and A. S. Perelson, Different dynamics of CD4+ and CD8+ T cell responses during and after acute lymphocytic choriomeningitis virus infection, J. Immunol., 171(2003), 3928–3935. |
| [58] | G. T. Skalski and J. F. Gilliam, Functional responses with predator interference: viable alternatives to the Holling type II model, Ecology, 82(2001), 3083–3092. |
| [59] | H. Nawata, M. T. Chong, D. Bronzert, et al., Estradiol-independent growth of a subline of MCF-7 human breast cancer cells in culture, J. Biol. Chem., 256(1981), 6895–6902. |
| [60] | R. Clarke, N. Brünner, B. S. Katzenellenbogen, et al., Progression of human breast cancer cells from hormone-dependent to hormone-independent growth both in vitro and in vivo, Proc. Natl. Acad. Sci. USA, 86(1989), 3649–3653. |
| [61] | N. T. Telang, G. Li, M. Katdare, et al., The nutritional herb Epimedium grandiflorum inhibits the growth in a model for the Luminal A molecular subtype of breast cancer, Oncol. Lett., 13 (2017), 2477–2482. |
| [62] | T. A. Caragine, M. Imai, A. B. Frey, et al., Expression of rat complement control protein Crry on tumor cells inhibits rat natural killer cell–mediated cytotoxicity, Blood, 100 (2002), 3304–3310. |
| [63] | M. R. Müller, F. Grünebach, A. Nencioni, et al., Transfection of dendritic cells with RNA induces CD4-and CD8-mediated T cell immunity against breast carcinomas and reveals the immunodominance of presented T cell epitopes, J. Immunol., 170 (2003), 5892–5896. |
| [64] | J. Chen, E. Hui, T. Ip, et al., Dietary flaxseed enhances the inhibitory effect of tamoxifen on the growth of estrogen-dependent human breast cancer (mcf-7) in nude mice, Clin. Cancer Res., 10(2004), 7703–7711. |
| [65] | P. V. Sivakumar, R. Garcia, K. S. Waggie, et al., Comparison of vascular leak syndrome in mice treated with IL21 or IL2, Comparative Med., 63 (2013), 13–21. |
| [66] | C.Wu, T.Motosha, H.A.Abdel-Rahman, etal., Freeandprotein-boundplasmaestradiol-17during the menstrual cycle, J. Clin. Endocrinol. Metab., 43(1976), 436–445. |
| [67] | E. Nikolopoulou, L. R. Johnson, D. Harris, et al., Tumour-immune dynamics with an immune checkpoint inhibitor, Lett. Biomath., 5(2018), S137–S159. |
| [68] | P. Vacca, E. Munari, N. Tumino, et al., Human natural killer cells and other innate lymphoid cells in cancer: friends or foes? Immunol. Lett., 201(2018), 14–19. |
| [69] | A. Cerwenka, J. Kopitz, P. Schirmacher, et al., HMGB1: the metabolic weapon in the arsenal of NK cells, Mol. Cell. Oncol., 3(2016), e1175538. |
| [70] | I. J. Fidler, Metastasis: quantitative analysis of distribution and fate of tumor emboli labeled with 125i-5-iodo-2'-deoxyuridine, J. Natl. Cancer Inst., 4(1970), 773–782. |
| [71] | G. G. Page and S. Ben-Eliyahu, A role for NK cells in greater susceptibility of young rats to metastatic formation, Dev. Comp. Immunol., 23(1999), 87–96. |
| [72] | O. E. Franco, A. K. Shaw, D. W. Strand, et al., Cancer associated fibroblasts in cancer pathogenesis, Semin. Cell Dev. Bio., 21(2010), 33–39. |
| [73] | G. P. Dunn, A. T. Bruce, H. Ikeda, et al., Cancer immunoediting: from immunosurveillance to tumor escape, Nat. Immunol., 3(2002), 991. |
| [74] | D. Mittal, M. M. Gubin, R. D. Schreiber, et al., New insights into cancer immunoediting and its three component phases-elimination, equilibrium and escape, Curr. Opin. Immunol., 27(2014), 16–25. |
| [75] | J. Nowak, P. Juszczynski and K. Warzocha, The role of major histocompatibility complex polymorphisms in the incidence and outcome of non-Hodgkin lymphoma, Curr. Immunol. Rev., 5(2009), 300–310. |
| [76] | J. G. B. Alvarez, M. González-Cao, N. Karachaliou, et al., Advances in immunotherapy for treatment of lung cancer, Cancer Biol. Med., 12(2015), 209–222. |
| [77] | M. E. Dudley and S. A. Rosenberg, Adoptive-cell-transfer therapy for the treatment of patients with cancer, Nat. Rev. Cancer, 3(2003), 666–675. |
| [78] | M. Su, C. Huang and A. Dai, Immune checkpoint inhibitors: therapeutic tools for breast cancer, Asian Pac. J. Cancer Prev., 17 (2016), 905–910. |
| [79] | M. Ebbo, L. Gérard, S. Carpentier, et al., Low circulating natural killer cell counts are associated with severe disease in patients with common variable immunodeficiency, EBioMedicine., 6(2016), 222–230. |
| [80] | S. H. Jee, J. Y. Park, H. Kim, et al., White blood cell count and risk for all-cause, cardiovascular, and cancer mortality in a cohort of Koreans, Am. J. Epidemiol., 162 (2005), 1062–1069. |
| [81] | W. B. Kannel, K. Anderson and T. W. Wilson, White blood cell count and cardiovascular disease: insights from the Framingham Study, Jama, 267 (1992), 1253–1256. |
| [82] | K. L. Margolis, J. E. Manson, P. Greenland, et al., Leukocyte count as a predictor of cardiovascular events and mortality in postmenopausal women: the Women's Health Initiative Observational Study, Arch. Intern. Med., 165(2005), 500–508. |
| [83] | B. K. Duffy, H. S. Gurm, V. Rajagopal, et al., Usefulness of an elevated neutrophil to lymphocyte ratio in predicting long-term mortality after percutaneous coronary intervention, Am. J. Cardiol., 97 (2006), 993–996. |
| [84] | B. D. Horne, J. L. Anderson, J. M. John, et al., Which white blood cell subtypes predict increased cardiovascular risk? J. Am. Coll. Cardiol., 45(2005), 1638–1643. |
| [85] | R. N. O. Cobucci, H. Saconato, P. H. Lima, et al., Comparative incidence of cancer in HIV-AIDS patients and transplant recipients, Cancer Epidemiol., 36(2012), e69–e73. |
| [86] | C. Bodelon, M. M. Madeleine, L. F. Voigt, et al., Is the incidence of invasive vulvar cancer increasing in the United States? Cancer Causes Control, 20(2009), 1779–1782. |
| [87] | M. R. Shurin, Cancer as an immune-mediated disease, Immunotargets Ther., 1 (2012), 1–6. |
| [88] | J. S. Lawrence, Leukopenia: its mechanism and therapy, J. Chronic. Dis., 6(1957), 351–364. |
| [89] | P. Venigalla, B. Motwani, A. Nallari, et al., A patient on hydroxyurea for sickle cell disease who developed an opportunistic infection, Blood, 100(2002), 363–364. |
| [90] | M. Iwamuro, S. Tanaka, A. Bessho, et al., Two cases of primary small cell carcinoma of the stomach, Acta. Med. Okayama, 63(2009), 293–298. |
| [91] | A. O. O. Chan, I. O. L. Ng, C. M. Lam, et al., Cholestatic jaundice caused by sequential carbimazole and propylthiouracil treatment for thyrotoxicosis, Hong Kong Med. J., 9(2003), 377–380. |
| [92] | J. H. Goodchild and M. Glick, A different approach to medical risk assessment, Endod. Topics, 4(2003), 1–8. |
| [93] | S. E. Hankinson, Endogenous hormones and risk of breast cancer in postmenopausal women, Breast Dis., 24(2006), 3–15. |
| [94] | R. Kaaks, S. Rinaldi, T. J. Key, et al., Postmenopausal serum androgens, oestrogens and breast cancer risk: the European prospective investigation into cancer and nutrition, Endocr. Relat. Cancer, 12(2005), 1071–1082. |
| [95] | S. A. Missmer, A. H. Eliassen, R. L. Barbieri, et al., Endogenous estrogen, androgen, and progesterone concentrations and breast cancer risk among postmenopausal women, J. Natl. Cancer Inst., 96(2004), 1856–1865. |
| [96] | A. A. Arslan, R. E. Shore, Y. Afanasyeva, et al., Circulating estrogen metabolites and risk for breast cancer in premenopausal women, Cancer Epidemiol. Biomarkers Prev., 18(2009), 2273–2279. |
| [97] | S. B. Brown and S. E. Hankinson, Endogenous estrogens and the risk of breast, endometrial, and ovarian cancers, Steroids, 99(2015), 8–10. |
| [98] | L. C. Houghton, D. Ganmaa, P. S. Rosenberg, et al., Associations of breast cancer risk factors with premenopausal sex hormones in women with very low breast cancer risk, Int. J. Environ. Res. Public Health, 13(2016), 1066. |
| [99] | R. Kaaks, K. Tikk, D. Sookthai, et al., Premenopausal serum sex hormone levels in relation to breast cancer risk, overall and by hormone receptor status-results from the EPIC cohortk, Int. J. cancer, 134(2014), 1947–1957. |
| [100] | A. Diefenbach, E. R. Jensen, A. M. Jamieson, et al., Rae1 and H60 ligands of the NKG2D receptor stimulate tumour immunity, Nature, 413(2001), 165. |
| [101] | A. Iannello and D. H. Raulet, Cold Spring Harbor symposia on quantitative biology, 1st edition, Cold Spring Harbor Laboratory Press, New York, 2013. |
| [102] | M. B. Pampena and E. M. Levy, Natural killer cells as helper cells in dendritic cell cancer vaccines, Front. Immunol., 6 (2015), 1–8. |
| [103] | E. Vivier, S. Ugolini, D. Blaise, et al., Targeting natural killer cells and natural killer T cells in cancer, Nat. Rev. Immunol., 12(2012), 239–252. |
| [104] | G. Liu, X. Fan, Y. Cai, et al., Efficacy of dendritic cell-based immunotherapy produced from cord blood in vitro and in a humanized NSG mouse cancer model, Immunotherapy, 11(2019), 599–616. |
| [105] | M. Schnekenburger, M. Dicato and M. F. Diederich, Anticancer potential of naturally occurring immunoepigenetic modulators: A promising avenue? Cancer, 125(2019), 1612–1628. |
| [106] | X. Feng, L. Lu, K. Wang, et al., Low expression of CD80 predicts for poor prognosis in patients with gastric adenocarcinoma, Future Oncol., 15 (2019), 473–483. |
| [107] | X. Lai and A. Friedman, Combination therapy of cancer with cancer vaccine and immune checkpoint inhibitors: a mathematical model, PloS One, 12 (2017), e0178479. |
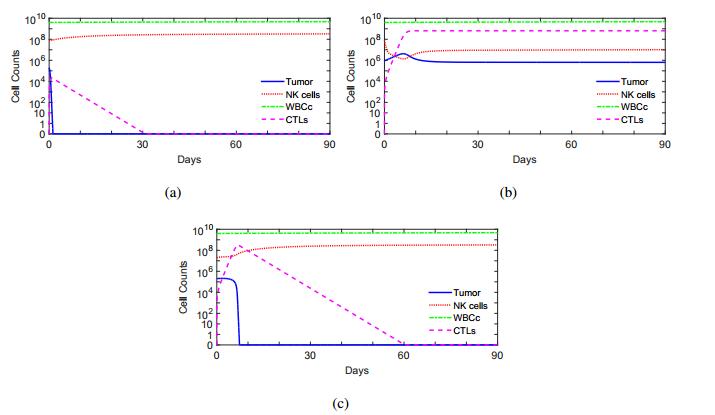
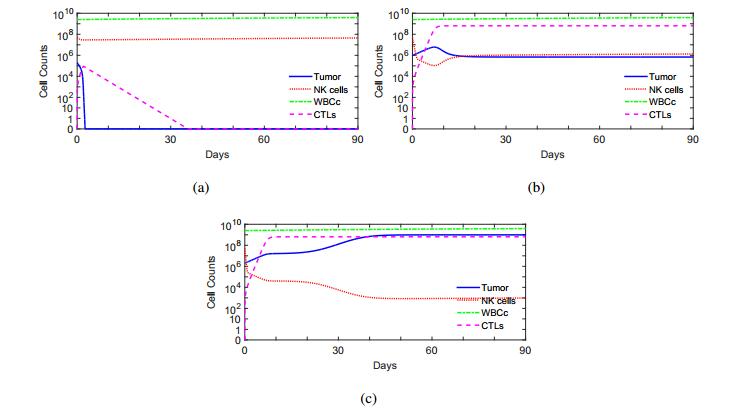
Figures(10) / Tables(1)

Hsiu-Chuan Wei. Mathematical modeling of tumor growth: the MCF-7 breast cancer cell line[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6512-6535. doi: 10.3934/mbe.2019325







 DownLoad:
DownLoad: