Citation: Sandesh Athni Hiremath, Christina Surulescu, Somayeh Jamali, Samantha Ames, Joachim W. Deitmer, Holger M. Becker. Modeling of pH regulation in tumor cells: Direct interaction between proton-coupled lactate transporters and cancer-associated carbonicanhydrase[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 320-337. doi: 10.3934/mbe.2019016

| [1] | J. Almquist, P. Lang, D. Prätzel-Wolters, J. W. Deitmer, M. Jirstrand and H. M. Becker, A Kinetic Model of the Monocarboxylate Transporter MCT1 and its Interaction with Carbonic Anhydrase II, J. Comp. Sci. Sys. Biol., 3 (2010), 107–116. |
| [2] | S. Ames, S. Pastorekova and H. M. Becker, The proteoglycan-like domain of carbonic anhydrase IX mediates non-catalytic facilitation of lactate transport in cancer cells, Oncotarget, 9 (2018), 27940–27957. |
| [3] | H. M. Becker and J. W. Deitmer, Nonenzymatic proton handling by carbonic anhydrase II during H+-lactate cotransport via monocarboxylate transporter 1, J. Biol. Chem., 283 (2008), 21655– 21667. |
| [4] | H. M. Becker, D. Hirnet, C. Fecher-Trost, D. Sültemeyer and J. W. Deitmer, Transport activity of MCT1 expressed in Xenopus oocytes is increased by interaction with carbonic anhydrase, J. Biol. Chem., 280 (2005), 39882–39889. |
| [5] | H. M. Becker, M. Klier and J. W. Deitmer, Nonenzymatic augmentation of lactate transport via monocarboxylate transporter isoform 4 by carbonic anhydrase II, J. Membrane Biol., 234 (2010), 125–135. |
| [6] | H. M. Becker, M. Klier, C. Schüler, R. McKenna and J. W. Deitmer, Intramolecular proton shuttle supports not only catalytic but also noncatalytic function of carbonic anhydrase II, P. Natl. Acad. Sci. USA., 108 (2011), 3071–3076. |
| [7] | M. C. Brahimi-Horn, G. Bellot and J. Pouysségur, Hypoxia and energetic tumour metabolism, Curr. Opin. Genet. Dev., 21 (2011), 67–72. |
| [8] | R. A. Cardone, V. Casavola and S. J. Reshkin, The role of disturbed pH dynamics and the Na+/H+ exchanger in metastasis, Nat. Rev. Cancer, 5 (2005), 786–795. |
| [9] | A. W. De Bruijne, H. Vreeburg and J. Van Steveninck, Kinetic analysis of L-lactate transport in human erythrocytes via the monocarboxylate-specific carrier system, Biochim. Biophys. Acta., 732 (1983), 562–568. |
| [10] | R. A. Gatenby and R. J. Gillies, Why do cancers have high aerobic glycolysis? Nat. Rev. Cancer, 4 (2004), 891–899. |
| [11] | A. P. Halestrap and N. T. Price, The proton-linked monocarboxylate transporter (MCT) family: structure, function and regulation Biochem. J., 343 (1999), 281–299. |
| [12] | D. Hanahan and R. A. Weinberg, Hallmarks of cancer: the next generation, Cell, 144 (2011), 646–674. |
| [13] | S. A. Hiremath, Multiscale modeling of acid mediated cancer invasion with random dynamics and beyond, Ph.D thesis, University of Kaiserslautern (TUK), 2017. |
| [14] | S. A. Hiremath, S. Sonner, C. Surulescu and A. Zhigun, On a coupled SDE-PDE system modeling acid-mediated tumor invasion, Discrete. Cont. Dyn.-B , 23 (2018), 2339–2369. |
| [15] | S. A. Hiremath and C. Surulescu, A stochastic multiscale model for acid mediated cancer invasion, Nonlinear. Anal., 22 (2015), 176–205. |
| [16] | S. A. Hiremath and C. Surulescu, A stochastic model featuring acid-induced gaps during tumor progression, Nonlinearity, 29 (2016), 851–914. |
| [17] | A. Innocenti, S. Pastorekova, J. Pastorek, A. Scozzafava, G. De Simone and C. T. Supuran, The proteoglycan region of the tumor-associated carbonic anhydrase isoform IX acts as anintrinsic buffer optimizing CO2 hydration at acidic pH values characteristic of solid tumors, Bioorg. Med. Chem. Lett., 19 (2009), 5825–5828. |
| [18] | M. Irving, J. Maylie, L. N. Sizto and K. W. Chandler, Intracellular diffusion in the presence of mobile buffers. Application to proton movement in muscle, Biophys. J., 57 (1990), 717–721. |
| [19] | S. Jamali, M. Klier, S. Ames, L. F. Barros, R. McKenna, J.W. Deitmer and H. M. Becker, Hypoxiainduced carbonic anhydrase IX facilitates lactate flux in human breast cancer cells by non-catalytic function, Sci. Rep., 5 (2015), 13605. |
| [20] | C. Juel A. P. and Halestrap, Lactate transport in skeletal muscle - role and regulation of the monocarboxylate transporter, J. Physiol., 517 (1999), 633–642. |
| [21] | P. E. Kloeden, S. Sonner and C. Surulescu, A nonlocal sample dependence SDE-PDE system modeling proton dynamics in a tumor, Discrete. Cont. Dyn.-B , 21 (2016), 2233–2254. |
| [22] | A. H. Lee and I. F. Tannock, Heterogeneity of intracellular pH and of mechanisms that regulate intracellular pH in populations of cultured cells, Cancer. Res., 58 (1998), 1901–1908. |
| [23] | C. Martin, S. F. Pedersen, A. Schwab and C. Stock, Intracellular pH gradients in migrating cells, Am. J. Physiol. Cell. Physiol., 300 (2011), C490–495. |
| [24] | C. Martinez, D. Kalise and L. F. Barros, General Requirement for Harvesting Antennae at Ca(2+) and H(+) Channels and Transporters, Front. Neuroenergetics, 2 (2010), 27. |
| [25] | P. E. Morgan, S. Pastorekova, A. K. Stuart-Tilley, S. L. Alper and J.R. Casey, Interactions of transmembrane carbonic anhydrase, CAIX, with bicarbonate transporters, Am. J. Physiol. Cell. Physiol., 293 (2007), C738–748. |
| [26] | S. I. Noor, S. Dietz, H. Heidtmann, C. D. Boone, R. McKenna, J. W. Deitmer and H.M. Becker, Analysis of the Binding Moiety Mediating the Interaction between Monocarboxylate Transporters and Carbonic Anhydrase II, J. Biol. Chem., 290 (2015), 4476–4486. |
| [27] | S. I. Noor, S. Jamali, S. Ames, S. Langer, J. W. Deitmer and H. M. Becker, A surface proton antenna in carbonic anhydrase II supports lactate transport in cancer cells, eLife 7 (2018), 1–31. |
| [28] | S. K. Parks, J. Chiche and J. Pouyssegur, pH control mechanisms of tumor survival and growth, J. Cell. Physiol., 226 (2011), 299–308. |
| [29] | S. K. Parks, J. Chiche and J. Pouyssegur, Disrupting proton dynamics and energy metabolism for cancer therapy, Nat. Rev. Cancer., 13 (2013), 611–623. |
| [30] | R. C. Poole and A. P. Halestrap, Transport of lactate and other monocarboxylates across mammalian plasma membranes, Am. J. Physiol., 264 (1993), C761–782. |
| [31] | P. E. Porporato, S. Dhup, R. K. Dadhich, T. Copetti and P. Sonveaux, Anticancer targets in the glycolytic metabolism of tumors: a comprehensive review, Front. Pharmacol., 2 (2011), 49. |
| [32] | A. Roos and W. F. Boron, Intracellular pH, Physiol. Rev., 61 (1981), 296–434. |
| [33] | A. Schulze and A. L. Harris, How cancer metabolism is tuned for proliferation and vulnerable to disruption, Nature, 491 (2012), 364–373. |
| [34] | O. Sedlakova, E. Svastova, M. Takacova, J. Kopacek, J. Pastorek and S. Pastorekova, Carbonic anhydrase IX, a hypoxia-induced catalytic component of the pH regulating machinery in tumors, Front. Physiol., 4 (2014), 400. |
| [35] | C. Stock and A. Schwab, Protons make tumor cells move like clockwork, Pflügers Archiv : European journal of physiology, 458 (2009), 981–992. |
| [36] | M. H. Stridh, M. D. Alt, S. Wittmann, H. Heidtmann, M. Aggarwal, B. Riederer, U. Seidler, G. Wennemuth, R. McKenna, J. W. Deitmer and H.M. Becker, Lactate flux in astrocytes is enhanced by a non-catalytic action of carbonic anhydrase II, J. Physiol., 590 (2012), 2333–2351. |
| [37] | E. Svastova, W. Witarski, L. Csaderova, T. Kosik, L. Skvarkova, A. Hulikova, M. Zatovicova, M. Barathova, J. Kopacek, J. Pastorek and S. Pastorekova, Carbonic anhydrase IX interacts with bicarbonate transporters in lamellipodia and increases cell migration via its catalytic domain, J. Biol. Chem., 287 (2012), 3392–3402. |
| [38] | P. Swietach, A. Hulikova, R. D. Vaughan-Jones and A.L. Harris, New insights into the physiological role of carbonic anhydrase IX in tumour pH regulation, Oncogene, 29 (2010), 6509– 6521. |
| [39] | P. Swietach, R. D. Vaughan-Jones, A. L. Harris and A. Hulikova, The chemistry, physiology and pathology of pH in cancer, Philosophical Transactions of the Royal Society of London. Series B, Biological sciences, 369 (2014), 20130099. |
| [40] | M. S. Ullah, A. J. Davies and A. P. Halestrap, The plasma membrane lactate transporter MCT4, but not MCT1, is up-regulated by hypoxia through a HIF-1alpha-dependent mechanism, J. Biol. Chem., 281 (2006), 9030–9037. |
| [41] | O. Warburg, F. Wind and E. Negelein, The metabolism of tumors in the body, J. Gen. Physiol., 8 (1927), 519–530. |
| [42] | S. Weinhouse, O. Warburg, D. Burk and A. L. Schade, On respiratory impairment in cancer cells, Science, 124 (1956), 269–270. |
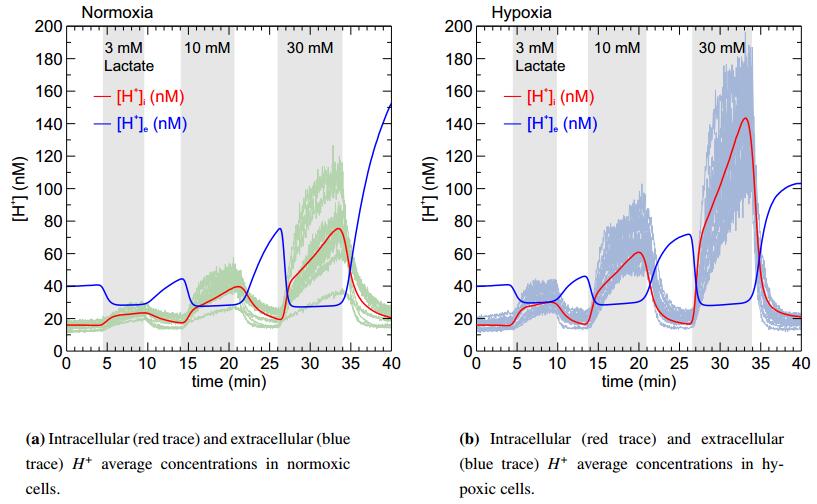
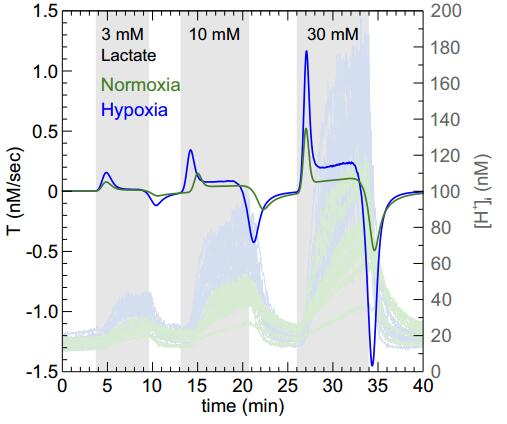
Figures(6)

Sandesh Athni Hiremath, Christina Surulescu, Somayeh Jamali, Samantha Ames, Joachim W. Deitmer, Holger M. Becker. Modeling of pH regulation in tumor cells: Direct interaction between proton-coupled lactate transporters and cancer-associated carbonicanhydrase[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 320-337. doi: 10.3934/mbe.2019016







 DownLoad:
DownLoad: