Citation: Ann Nwankwo, Daniel Okuonghae. Mathematical assessment of the impact of different microclimate conditions on malaria transmission dynamics[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1414-1444. doi: 10.3934/mbe.2019069

| [1] | G. J. Abiodun, R. Maharaj, P. Witbooi, et al., Modelling the influence of temperature and rainfall on the population dynamics of anopheles arabiensis, Malaria J., 15 (2016), 364. |
| [2] | Y. A. Afrane, B.W. Lawson, A. K. Githeko, et al., Effects of microclimatic changes caused by land use and land cover on duration of gonotrophic cycles of anopheles gambiae (Diptera: Culicidae) in western Kenya highlands, J. Med. Entomol., 42 (2005), 974–980. |
| [3] | F. B. Agusto, A. B. Gumel and P. E. Parham, Qualitative assessment of the role of temperature variation on malaria transmission dynamics, J. Biol. Syst., 24 (2015), 1–34. |
| [4] | N. Bacaer, Approximation of the basic reproduction number Ro for vector-borne diseases with a periodic vector population, B. Math. Biol., 69 (2007), 1067–1091. |
| [5] | L. M. Beck-Johnson, W. A. Nelson, K. P. Paaijmans, et al., The importance of temperature fluctuations in understanding mosquito population dynamics and malaria risk, Roy. Soc. Open Sci., 4 (2017). Available from :http://dx.doi.org/10.1098/rsos.160969. |
| [6] | J. I. Blanford, S. Blanford, R. G. Crane, et al., Implications of temperature variation for malaria parasite development across Africa, Sci. Rep., 3 (2013), 1300. |
| [7] | P. Cailly, A. Tran, T. Balenghien, et al., A climate-driven abundance model to assess mosquito control strategies, Ecol. Model., 227 (2012), 7–17. |
| [8] | C. Christiansen-Jucht, K. Erguler, C. Y. Shek, et al., Modelling anopheles gambiae s.s. population dynamics with temperature and age-dependent survival, Int. J. Env. Res. Pub. He., 12 (2015), 5975–6005. |
| [9] | O. Diekmann, J. Heesterbeek and J. Metz, On the definition and the computation of the basic reproduction ratio R0 in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28 (1990), 365–382. |
| [10] | Y. Dumont and F. Chiroleu, Vector control for the chikungunya disease, Math. Biosci. Eng., 7 (2010), 105–111. |
| [11] | A. Egbendewe-Mondzozo, M. Musumba, B. A. McCarl, et al., Climate change and vector-borne diseases: an economic impact analysis of malaria in Africa, Int. J. Env. Res. Pub. He., 8 (2011), 913–930. |
| [12] | S. M. Garba, A. B. Gumel and M. R. A. Bakar, Backward bifurcations in dengue transmission dynamics, Math. Biosci., 215 (2008), 11–25. |
| [13] | A. B. Gumel, Causes of backward bifurcation in some epidemiological models, J. Math. Anal. Appl., 395 (2012), 355–365. |
| [14] | V. Lakshmikantham and S. Leela, Differential and Integral Inequalities: Theory and Applications, Academic Press, New York, 1969. |
| [15] | J. La Salle and S. Lefschetz, The stability of dynamical systems, SIAM, Philadephia, 1976. |
| [16] | Y. Lou and X. Q. Zhao, A climate-based malaria transmission model with structured vector population, SIAM J. Appl. Math., 70 (2010), 2023–2044. |
| [17] | P. Magal and X. Q. Zhao, Global attractors and steady states for uniformly persistent dynamical systems, SIAM J. Math. Anal., 37 (2005), 251–275. |
| [18] | E. A. Mordecai, Optimal temperature for malaria transmission is dramatically lower than previously predicted, Ecol. Lett., 16 (2013), 22–30. |
| [19] | E. T. Ngarakana-Gwasira, C. P. Bhunu, M. Masocha, et al., Assessing the role of climate change in malaria transmission in Africa, Malaria Res. Treat., 1 (2016), 1–7. |
| [20] | C. N. Ngonghala, S. Y. Del Valle, R. Zhao, et al., Quantifying the impact of decay in bed-net efficacy on malaria transmission, J. Theor. Biol., 363 (2014), 247–261. |
| [21] | H. S. Ngowo, E. W. Kaindoa, J. Matthiopoulos, et al., Variations in household microclimate affect outdoor-biting behaviour of malaria vectors, Wellcome Open Research, 2 (2017). |
| [22] | A. M. Niger and A. B. Gumel, Mathematical analysis of the role of repeated exposure on malaria transmission dynamics, Diff. Equa. Dyn. Syst., 16 (2008), 251–287. |
| [23] | K. Okuneye and A. B. Gumel, Analysis of a temperature- and rainfall-dependent model for malaria transmission dynamics, Math. Biosci., 287 (2017), 72–92. |
| [24] | K. P. Paaijmans, A. F. Read and M. B. Thomas, Understanding the link between malaria risk and climate, P. Natl. Acad. Sci. USA, 106 (2009), 13844–13849. |
| [25] | K. P. Paaijmans, S. Blanford, A. S. Bell, et al., Influence of climate on malaria transmission depends on daily temperature variation, P. Natl. Acad. Sci. USA, 107 (2010), 15135–15139. |
| [26] | K. P. Paaijmans, S. S. Imbahale, M. B. Thomas, et al., Relevant microclimate for determining thedevelopment rate of malaria mosquitoes and possible implications of climate change, Malaria J., 9 (2010), 196. |
| [27] | K. P. Paaijmans and M. B. Thomas, The influence of mosquito resting behaviour and associated microclimate for malaria risk, Malaria J., 10 (2011), 183. |
| [28] | K. P. Paaijmans and M. B. Thomas, Relevant temperatures in mosquito and malaria biology, In: Ecology of parasite-vector interactions, Wageningen Academic Publishers, 2013. |
| [29] | P. E. Parham and E. Michael, Modeling the effects of weather and climate change on malaria transmission, Environ. Health Persp., 118 (2010), 620–626. |
| [30] | D. J. Rogers and S. E. Randolph, Advances in Parasitology, Elsevier Academic Inc, San Diego, 2006. |
| [31] | M. A. Safi, M. Imran and A. B. Gumel, Threshold dynamics of a non-autonomous SEIRS model with quarantine and isolation, Theor. Biosci., 131 (2012), 19–30. |
| [32] | L. L. M. Shapiro, S. A. Whitehead and M. B. Thomas, Quantifying the effects of temperature on mosquito and parasite traits that determine the transmission potential of human malaria, PLoS Biol., 15 (2017), e2003489. |
| [33] | P. Singh, Y. Yadav, S. Saraswat, et al., Intricacies of using temperature of different niches for assessing impact on malaria transmission, Indian J. Med. Res., 144 (2016), 67–75. |
| [34] | H. L. Smith, Monotone dynamical systems: an introduction to the theory of competitive and cooperative systems, Am. Math. Soc., 41, 1995. |
| [35] | S. Thomas, S. Ravishankaran, N. A. J. Amala Justin, et al., Microclimate variables of the ambient environment deliver the actual estimates of the extrinsic incubation period of plasmodium vivax and plasmodium falciparum: a study from a malaria endemic urban setting, Chennai in India, Malaria J., 17 (2018), 201. |
| [36] | P. Van den Driessche and J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
| [37] | W. Wang and X. Q. Zhao, Threshold dynamics for compartmental epidemic models in periodic environments, J. Dyn. Differ. Equations, 20 (2008), 699–717. |
| [38] | Malaria Report, from World Health Organization, 2010. Available from: www.who.int/ mediacenter/factsheets/fs094/en/. |
| [39] | World Malaria Day, Report of the World Health Organisation (WHO), 2018. Available from: www.who.int/malaria/media/world-malaria-day-2018/en/. |
| [40] | H. Zhang, P. Georgescu and A. S. Hassan, Mathematical insights and integrated strategies for the control of Aedes aegypti mosquito, Appl. Math. Comput., 273 (2016), 1059–1089. |
| [41] | F. Zhang and X. Q. Zhao, A periodic epidemic model in a patchy environment, J. Math. Anal. Appl., 325 (2007), 496–516. |
| [42] | X. Q. Zhao, Dynamical systems in population biology, Springer, New York, 2003. |
| [43] | X. Q. Zhao, Uniform persistence and periodic co-existence states in infinite-dimensional periodic semiflows with applications, Can. Appl. Math. Q., 3 (1995), 473–495. |
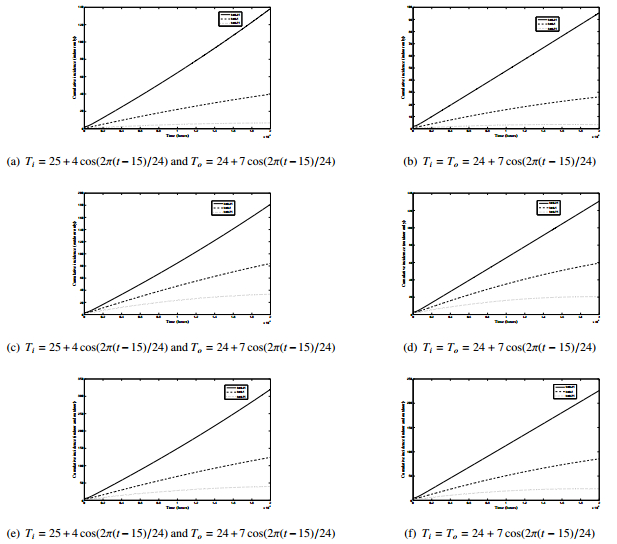
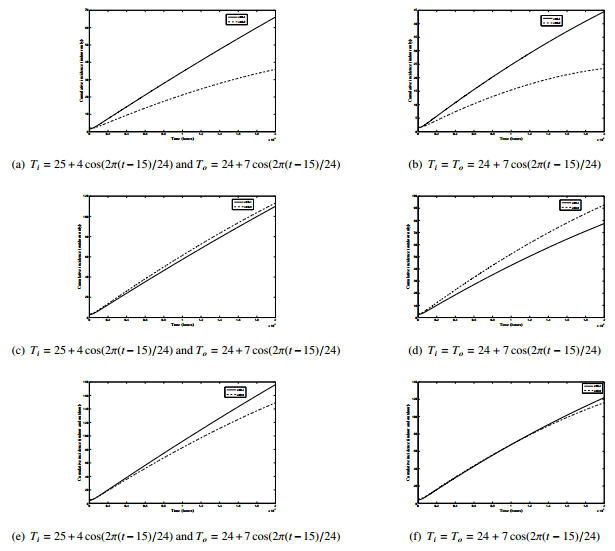
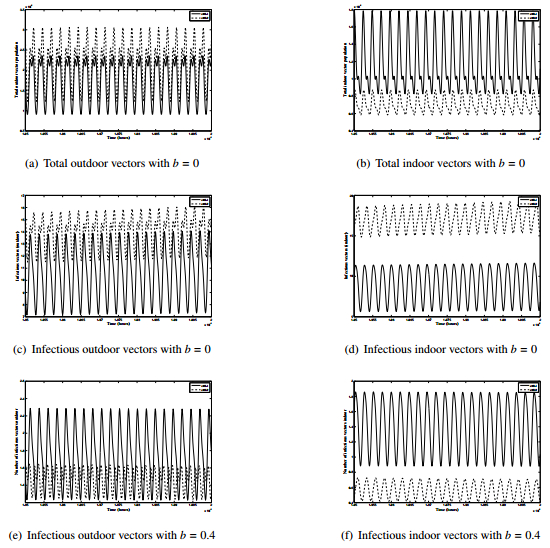
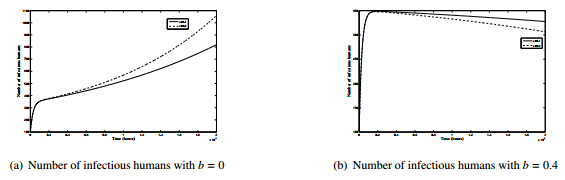
Figures(8) / Tables(2)

Ann Nwankwo, Daniel Okuonghae. Mathematical assessment of the impact of different microclimate conditions on malaria transmission dynamics[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1414-1444. doi: 10.3934/mbe.2019069







 DownLoad:
DownLoad: