Citation: Ivan Nygaard, Famakan Kamissoko, Per Bromand Nørgård, Jake Badger, Luc Dewilde. Feasibility of wind power integration in weak grids in non-coastal areas of sub-saharan Africa: the case of Mali[J]. AIMS Energy, 2017, 5(3): 557-584. doi: 10.3934/energy.2017.3.557

| [1] | UN (2012) International Year of Sustainable Energy for All, 2012, Report of the Secretary-General; United Nations General Assembly, Sixty-seventh session, Item 20 of the provisional agenda. Available from: http://www.se4all.org/sites/default/files/l/2015/08/A-67-314-2012-Report-of-the-Secretary-General-to-the-General-Assembly-on-the-Year-of-Sustainable-Energy-for-All.pdf. |
| [2] | IEA (2014) World Energy Outlook 2014 Electricity Access Database Available from: http://www.worldenergyoutlook.org/resources/energydevelopment/energyaccessdatabase/. |
| [3] | Eberhard A, Foster V, Briceño GC, et al. (2010) Catching Up. In: Foster V, Briceño-Garmendia C, Africa's Infrastructure: A time for transformation; IBRD/WB: Washington DC, 181–202. |
| [4] |
Bazilian M, Nussbaumer P, Rogner HH, et al. (2012) Energy access scenarios to 2030 for the power sector in sub-Saharan Africa. Util Policy 20: 1–16. doi: 10.1016/j.jup.2011.11.002

|
| [5] | IRENA (2012) Prospects for the African Power Sector; International Renewable Energy Agency (IRENA), Abu Dhabi, United Arab Emirates. Available from: https://www.irena.org/DocumentDownloads/Publications/Prospects_for_the_African_PowerSector.pdf. |
| [6] | GWEC (2016) Global wind energy outlook; Global Wind Energy Council (GWEC). Available from: http://www.gwec.net/publications/global-wind-energy-outlook/global-wind-energy-outlook-2016/. |
| [7] | Tarnoy H (2014) Lake Turkana Wind Power Project (LTWP); Presentation at Seminar on Sustainable Energy Investments in Africa-Engaging the Private Sector UN City, Copenhagen, 25 June 2014. Available from: http://www.unepdtu.org/PUBLICATIONS/Workshop-Presentations/Workshop-Presentations-Sustainable-Energy-Investments-in-Africa. |
| [8] |
Msimanga B, Sebitosi AB (2014) South Africa's non-policy driven options for renewable energy development. Renew Energ 69: 420–427. doi: 10.1016/j.renene.2014.03.041
|
| [9] | Alhamwi A, Kleinhans D, Weitemeyer S, et al. (2015) Moroccan national energy strategy reviewed from a meteorological perspective. Energy Strateg Rev 6: 39–47. |
| [10] | GWEC (2014) Global Wind Energy Outlook 2014; Global Wind Energy Council (GWEC). Available from: http://www.gwec.net/publications/global-wind-energy-outlook/global-wind-energy-outlook-2016/. |
| [11] | Adaramola MS, Agelin CM, Paul SS (2014) Assessment of wind power generation along the coast of Ghana. Energ Convers Manage 77: 61–69. |
| [12] | Ahiataku TW (2014) Perspectives on Sustainable Energy Investment in Ghana; Presentation at Seminar on Sustainable Energy Investments in Africa-Engaging the Private Sector UN City, Copenhagen, 25 June 2014. Available from: http://www.unepdtu.org/PUBLICATIONS/ Workshop-Presentations/Workshop-Presentations---Sustainable-Energy-Investments-in-Africa. |
| [13] | Dodd J (2014) West Africa's largest project moves closer; Wind Power Monthly, 6 January 2014. Available from: http://www.windpowermonthly.com/article/1226020/west-africas-largest-project-moves-closer. |
| [14] |
Mentis D, Hermann S, Howells M, et al. (2015) Assessing the technical wind energy potential in Africa a GIS-based approach. Renew Energ 83: 110–125. doi: 10.1016/j.renene.2015.03.072
|
| [15] |
Kousksou T, Allouhi A, Belattar M, et al. (2015) Renewable energy potential and national policy directions for sustainable development in Morocco. Renew Sust Energ Rev 47: 46–57. doi: 10.1016/j.rser.2015.02.056
|
| [16] |
El Alimi S, Maatallah T, Dahmouni A, et al. (2012) Modeling and investigation of the wind resource in the gulf of Tunis, Tunisia. Renew Sust Energ Rev 16: 5466–5478. doi: 10.1016/j.rser.2012.05.004
|
| [17] | Ahmed AS (2012) Potential wind power generation in South Egypt. Renew Sust Energ Rev 16: 1528–1536. |
| [18] | Mortensen NG, Said SU, Badger J (2006). Wind Atlas for Egypt. In Proceedings of the Third Middle East-North Africa Renewable Energy Conference (on CD-ROM). Available from: http://orbit.dtu.dk/files/52612711/Wind_Atlas_for_Egypt_paper.pdf. |
| [19] |
Asress MB, Simonovic A, Komarov D, et al. (2013) Wind energy resource development in Ethiopia as an alternative energy future beyond the dominant hydropower. Renew Sust Energ Rev 23: 366–378. doi: 10.1016/j.rser.2013.02.047
|
| [20] |
Ayodele TR, Jimoh AA, Munda J, et al. (2012) Wind distribution and capacity factor estimation for wind turbines in the coastal region of South Africa. Energ Convers Manage 64: 614–625. doi: 10.1016/j.enconman.2012.06.007
|
| [21] |
Bilal BO, Ndongo M, Kebe CMF, et al. (2013) Feasibility study of wind energy potential for electricity generation in the northwestern coast of senegal. Energ Procedia 36: 1119–1129. doi: 10.1016/j.egypro.2013.07.127
|
| [22] |
Boudia SM, Benmansour A, Ghellai N, et al. (2013) Temporal assessment of wind energy resource at four locations in Algerian Sahara. Energ Convers Manage 76: 654–664. doi: 10.1016/j.enconman.2013.07.086
|
| [23] |
Kamau JN, Kinyua R, Gathua JK (2010) 6 years of wind data for Marsabit, Kenya average over 14 m/s at 100 m hub height; An analysis of the wind energy potential. Renew Energ 35: 1298–1302. doi: 10.1016/j.renene.2009.10.008
|
| [24] |
Mukulo BM, Ngaruiya JM, Kamau JN (2014) Determination of wind energy potential in the Mwingi-Kitui plateau of Kenya. Renew Energ 63: 18–22. doi: 10.1016/j.renene.2013.08.042
|
| [25] | Badger J, Frank H, Hahmann AN, et al. (2014) Wind-climate estimation based on mesoscale and microscale modeling: statistical–dynamical downscaling for wind energy applications. J Appl Meteorol Clim 53: 1901–1919. |
| [26] | Nygaard I, Nørgård P, Dewilde L, et al. (2012) Screening of feasible applications of wind and solar energy in Mali: Assessment using the solar and wind atlas for Mali; UNEP Risoe Centre, Technical University of Denmark. Available from: http://www.frsemali.org/reports/00%20final%20reports/Screening%20of%20applications.pdf . |
| [27] | Badger J, Kamissoko F, Rasmussen MO, et al. (2012) Estimation of wind and solar resources in Mali; UNEP Risø Centre, Technical University of Denmark. Available from: http://www.frsemali.org/reports/00%20final%20reports/Estimation%20of%20solar%20and%20wind%20resources.pdf. |
| [28] | de Volder G, Dewilde L, Diallo M, et al. (2009) Systéme éolien-diesel á Tombouctou: etude de faisabilité; Coopération technique Belge, CNESOLER, Mali. |
| [29] | Karhammer R, Sanghvi A, Fernstrom E, et al. (2006) Sub-Saharan Africa: Introducing Low Cost Methods in Electricity Distribution Networks; ESMAP technical paper 104/06. Available from: http://documents.worldbank.org/curated/en/home. |
| [30] | SOGREAH (2009) Etude de la demande et du plan directeur d'investissement optimaux dans le secteur de l'électricité au Mali: Synthese et recommenda-tions; Ministére de l'Energie, des Mines et de l'Eau - Direction Nationale de l'Energie et SOGREAH. |
| [31] | GTZ (2004) Feasibility Study for a 900-kW Wind Farm in Gao, Mali. Wind-Diesel systm. Final report; Deutsche Gesellschaft für Technische Zusammenarbeit (GTZ) GmbH: Available from: http://www.gtz.de/de/dokumente/en-windenergy-mali-feasibility-study-gao-2004.pdf. |
| [32] | Cold-Ravnkilde SM (2013) War and Peace in Mali: Background and perspectives; Danish Institute for International Studies, Denmark. |
| [33] |
Eberhard A, Kåberger T (2016) Renewable energy auctions in South Africa outshine feed-in tariffs. Energ Sci Eng 4: 190–193. doi: 10.1002/ese3.118
|
| [34] | Nygaard I, Rasmussen K, Badger J, et al. (2010) Using modeling, satellite images and existing global datasets for rapid preliminary assessments of renewable energy resources: the case of Mali. Renew Sust Energ Rev 14: 2359–2371. |
| [35] | Pueyo A, Bawakyillenuo S, Osiolo H (2016) Cost and Returns on Renewable Energy in Sub-Saharan Africa: A comparison of Kenya and Ghana; Institute of Development Studiess, UK. Available from: https://opendocs.ids.ac.uk/opendocs/bitstream/handle/123456789/11297/ER190_ CostandReturnsofRenewableEnergyinSubSaharanAfricaAComparisonofKenyaandGhana.pdf;jsessionid=F5B7F647B177FA09989CD5399849283E?sequence=1. |
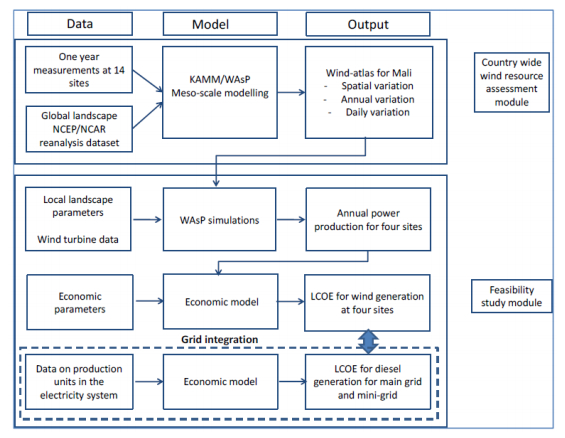
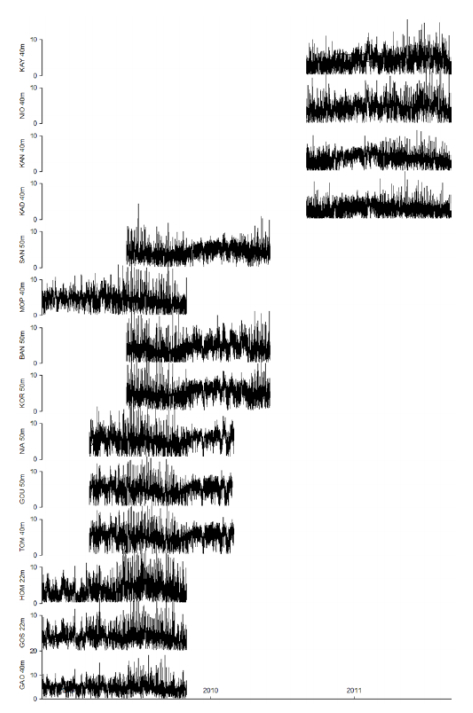

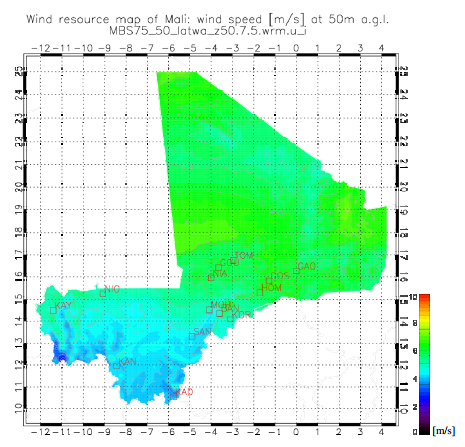
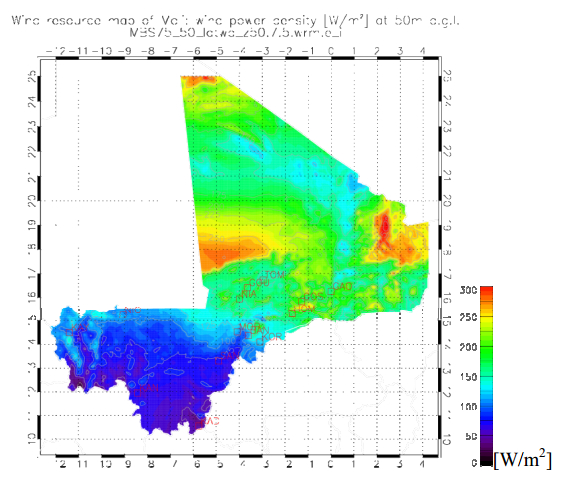
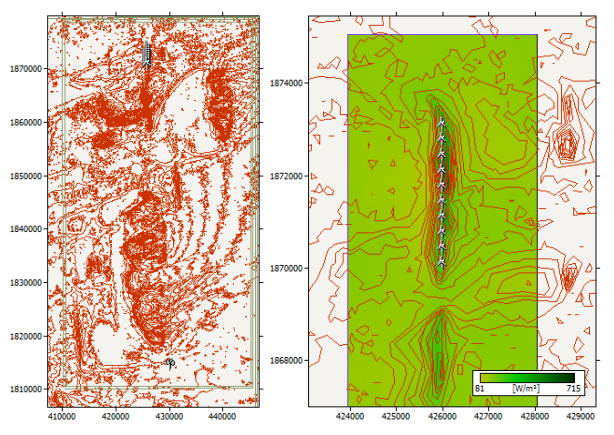
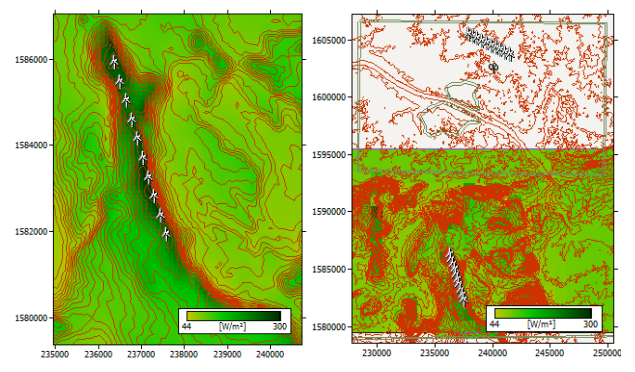
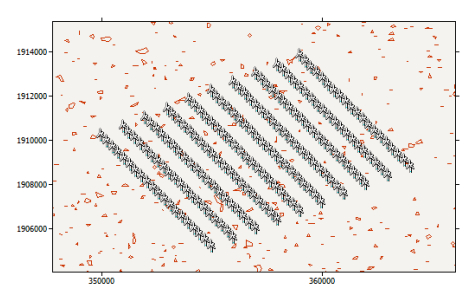
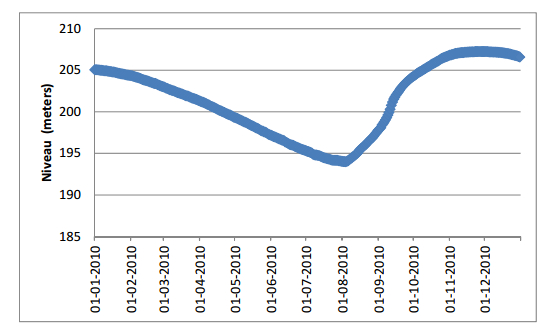
Figures(15) / Tables(8)

Ivan Nygaard, Famakan Kamissoko, Per Bromand Nørgård, Jake Badger, Luc Dewilde. Feasibility of wind power integration in weak grids in non-coastal areas of sub-saharan Africa: the case of Mali[J]. AIMS Energy, 2017, 5(3): 557-584. doi: 10.3934/energy.2017.3.557







 DownLoad:
DownLoad: