Citation: Ernest Greene, Michael J. Hautus. Demonstrating Invariant Encoding of Shapes Using A Matching Judgment Protocol[J]. AIMS Neuroscience, 2017, 4(3): 120-147. doi: 10.3934/Neuroscience.2017.3.120

| [1] | Kohler W (1938) Gestaltprobleme und Anfänge einer Gestalttheorie. Jahresbericht über der Gesellshaft Physiologie. Translated by: Ellis WD, A Source Book of Gestalt Psychology, London: Routledge & Kegan Paul: 58. |
| [2] | Goldmeier E (1936/1972) Similarity in visually perceived forms. Psychol Issues 8: 1-135. |
| [3] | Greene E (2007) Recognition of objects displayed with incomplete sets of discrete boundary dots. Percept Mot Skills 104: 1043-1059. |
| [4] |
Greene E, Visani A (2015) Recognition of letters displayed as briefly flashed dot patterns. Atten Percept Psychophys 77: 1955-1969. doi: 10.3758/s13414-015-0913-6

|
| [5] | Greene E (2016) Information persistence evaluated with low-density dot patterns. Acta Psychol 170: 215-225. |
| [6] | Green DM, Swets JA (1966) Signal Detection Theory and Psychophysics. New York; Wiley. |
| [7] |
Hautus MJ, van Hout D, Lee HS (2009) Variants of A Not-A and 2AFC tests: Signal Detection Theory models. Food Qual Prefer 20: 222-229. doi: 10.1016/j.foodqual.2008.10.002
|
| [8] | Macmillan NA, Creelman CD (2005) Detection Theory: A User's Guide, New Jersey: Lawrence Erlbaum. |
| [9] |
Hautus MJ (1995) Corrections for extreme proportions and their biasing effects on estimated values of d´. Behav Res Methods Instrum Comput 27: 46-51. doi: 10.3758/BF03203619
|
| [10] |
Miller J (1996) The sampling distribution of d'. Percept Psychophys 58: 65-72. doi: 10.3758/BF03205476
|
| [11] | Hautus J (2012) SDT Assistant (version 1.0) [Software]. Available from http://hautus.org. |
| [12] | Hautus MJ (1997) Calculating estimates of sensitivity from group data: pooled versus averaged estimators. Behav Res Methods Instrum Comput 29: 556-562. |
| [13] | Hubel DH, Wiesel TN (1959) Receptive fields of single neurons in the cat's striate cortex. J Physiol 148: 574-591. |
| [14] | Hubel DH, Wiesel TN (1968) Receptive fields and functional architecture of monkey striate cortex. J Physiol 195: 215-243. |
| [15] | Selfridge OG (1959) Pandemonium: A Paradigm for Learning in the Mechanization of Thought Process, London; HM Stationary Office. |
| [16] |
Sutherland NS (1968) Outlines of a theory of visual pattern recognition in animals and man. Proc R Soc Lond B Biol Sci 171: 297-317. doi: 10.1098/rspb.1968.0072
|
| [17] | Binford TO (1971) Visual perception by computer. Proc IEEE Conf Syst Control, Miami, FL. |
| [18] |
Barlow HB (1972) Single units and sensation: a neuron doctrine for perceptual psychology. Perception 1: 371-394. doi: 10.1068/p010371
|
| [19] | Milner PM (1974) A model for visual shape recognition. Psychol Rev 81: 521-535. |
| [20] | Palmer SE (1975) Visual perception and world knowledge: notes on a model of sensory-cognitive interaction. In: Norman DA, Rumelhart DE eds, Explorations in Cognitions, San Francisco: WH Freeman & Co.: 279-307. |
| [21] | Marr D (1982) Vision: A Computational Investigation into the Human Representation and Processing of Information, New York: Freemen: 51-79. |
| [22] | DeValois RL, Devalois KK (1991) Vernier acuity with stationary moving Gabors. Vision Res 31: 1619-1626. |
| [23] |
Panda R, Chatterji BN (1996) Gabor function: an efficient tool for digital image processing. IETE Tech Rev 13: 225-231. doi: 10.1080/02564602.1996.11416611
|
| [24] | Kohonen T, Oja E (1998) Visual feature analysis by the self-organizing maps. Neural Comput Appl 7: 273-286. |
| [25] |
Pettet MW, McKee SP, Grzywacz NM (1998) Constraints on long range interactions mediating contour detection. Vision Res 38: 865-879. doi: 10.1016/S0042-6989(97)00238-1
|
| [26] |
Pennefather PM, Chandna A, Kovacs I, et al. (1999) Contour detection threshold: repeatabililty and learning with 'contour cards.' Spat Vis 12: 257-266. doi: 10.1163/156856899X00157
|
| [27] | Taylor G, Hipp D, Moser A, et al. (2014) The development of contour processing: evidence from physiology and psychophysics. Front Psychol 5: e719 |
| [28] | Dong X, Chantlier MJ (2016) Perceptually motivated image features using contours. IEEE Trans Image Process 25: 5050-5062. |
| [29] | Edelman S (1999) Representation and Recognition in Vision, MIT Press. |
| [30] |
Cooke T, Jakel F, Wallraven C, et al. (2007) Multimodal similarity and categorization of novel, three-dimensional objects. Neuropsychologia 45: 484-495. doi: 10.1016/j.neuropsychologia.2006.02.009
|
| [31] | Hayworth KJ (2012) Dynamically partitionable autoassociative networks as a solution to the neural binding problem. Front Comput Neurosci 6: e73. |
| [32] |
Rodriguez-Sanchez AJ, Tsotsos JK (2012) The roles of endstopped and curvature tuned computations in a hierarchical representation of 2D shape. PLoS ONE 7: e42058. doi: 10.1371/journal.pone.0042058
|
| [33] | Hopfield JJ (1995) Pattern recognition computation using action potential timing for stimulus representation. Nature 376: 33-36. |
| [34] | Hopfield JJ (1996) Transforming neural computations and representing time. Proc Nat Acad Sci 83: 15440-15444. |
| [35] |
Maass W (1997) Fast sigmoidal networks via spiking neurons. Neural Comput 9: 279-304. doi: 10.1162/neco.1997.9.2.279
|
| [36] | McClelland JL (2013) Integrating probabilistic models of perception and interactive neural networks: a historical and tutorial review. Front Psychol 4: e503. |
| [37] | Guan T, Wang Y, Duan L, et al. (2015) On-device mobile landmark recognition using binarized descriptor and multifeature fusion. ACM Trans Intell Syst Technol 7: e12. |
| [38] |
Wei B, Guan T, Duan L, et al. (2015) Wide area localization and tracking on camera phones for mobile segmented reality systems. Multimed Syst 21: 381-399. doi: 10.1007/s00530-014-0364-2
|
| [39] | Pan H, Guan T, Luo Y, et al. (2016) Dense 3D reconstruction combining depth and RGB information. Neurocomputing 175: 644-651. |
| [40] | Ullman S (1976) Filling-in the gaps: the shape of subjective contours and a model for their generation. Biol Cybern 25: 1-6. |
| [41] | Sha'ashua A, Ullman S (1988) Structural saliency: the detection of globally salient structures using a locally connected network. In: Proc 2nd Intern Conf Comput Vision, Clearwater FL: 321-327. |
| [42] |
Kellman PJ, Shipley TF (1991) A theory of visual interpolation in object rperception. Cogn Psychol 23: 141-221. doi: 10.1016/0010-0285(91)90009-D
|
| [43] | Shipley TF, Kellman PJ (1992) Strength of visual interpolation depends on the ratio of physically specified to total edge length. Percept Psychophys 52: 97-106. |
| [44] | Cormen TH, Leherson CE, Rivest RL, et al. (2001) Single-source shortest paths and all pairs shortest paths. In: Introduction to Algorithms, MIT Press & McGraw-Hill: 580-642. |
| [45] | Field DJ, Hayes A, Hess RF (1993) Contour integration by the human visual system: evidence for a local "association field." Vision Res 33: 173-193. |
| [46] | Kwon TK, Agrawal K, Li Y, et al. (2016) Spatially-global integration of closed, fragmented contours by finding the shortest-path in a log-polar representation. Vision Res 126: 143-163. |
| [47] | Sceniak MP, Hawken MJ, Shapley R (2001) Visual spatial characterization of Macaque V1 neurons. J Neurophysiol 85: 1873-1887. |
| [48] | Bowers JS (2009) On the biological plausibility of grandmother cells: implications for neural network theories in psychology and neuroscience. Psychol Rev 116: 220-251. |
| [49] | Greene E (2008) Additional evidence that contour attributes are not essential cues for object recognition. Behav Brain Funct 4: e26. |
| [50] | Greene E (2016) How do we know whether three dots form an equilateral triangle? JSM Brain Sci 1: 1002. |
| [51] |
Lichtsteiner P, Posch C, Delbruck T (2008) A 128x128 120 dB 15 µs latency asynchronous temporal contrast vision sensor. IEEE J Solid-State Circuits 43: 566-576. doi: 10.1109/JSSC.2007.914337
|
| [52] | Robinson DA (1964) The mechanisms of human saccadic eye movement. J Physiol 174: 245-264. |
| [53] | Zimmermann E, Lappe M (2016) Visual space constructed by saccadic motor maps. Front Human Neurosci 10: e225. |
| [54] |
McSorlely E, McCloy R, Williams L (2016) The concurrent programming of saccades. PLoS ONE 11: e0168724. doi: 10.1371/journal.pone.0168724
|
| [55] | Bhutani N, Sengupta S, Basu D, et al. (2017) Parallel activation of prospective motor plans during visually-guided sequential saccades. Neurosci 45: 631-642. |
| [56] |
Feldman JA, Ballard DH (1982) Connectionist models and their properties. Cogn Sci 6: 205-254. doi: 10.1207/s15516709cog0603_1
|
| [57] |
Fukushima K (1980) Neocognitron: a self-organizing neural network model for a mechanism of pattern recognition unaffected by shift in position. Biol Cybern 36: 193-202. doi: 10.1007/BF00344251
|
| [58] |
Rolls ET (1992) Neurophysiological mechanisms underlying face processing within and beyond the temporal cortical visual areas. Phil Trans R Soc 335: 11-21. doi: 10.1098/rstb.1992.0002
|
| [59] | Wallis G, Rolls ET (1997) Invariant face and object recognition in the visual system. Prog Neurobiol 51: 167-194. |
| [60] | Riesenhuber M, Poggio T (2000) Models of object recognition. Nature Neurosci (S3): 1199-1204. |
| [61] | Pasupathy A, Connor CE (2001) Shape representation in area V4: position-specific tuning for boundary conformation. J Neurophysiol 86: 2505-2519. |
| [62] |
Suzuki N, Hashimoto N, Kashimori Y, et al. (2004) A neural model of predictive recognition in form pathway of visual cortex. BioSystems 76: 33-42. doi: 10.1016/j.biosystems.2004.05.004
|
| [63] |
Pinto N, Cox DD, DeCarlo JJ (2008) Why is real-world visual object recognition hard? PLoS Comput Biol 4: e27. doi: 10.1371/journal.pcbi.0040027
|
| [64] | Hancock PJB, Walton L, Mitchell G, et al. (2008) Segregation by onset asynchrony. J Vision 8: 1-21. |
| [65] | Karplus I, Goren M, Algorn D (1982) A preliminary experimental analysis of predator face recognition by Chromis caenuleus (Pisces, Pomacentridae). Z Tierpsychol 58: 53-65. |
| [66] | Siebeck UE, Parker AN, Sprenger D, et al. (2010) A species of reef fish that uses untraviolet patterns for covert face recognition. Curr Biol 20: 407-410. |
| [67] | Karplus I, Katzenstein R, Goren M (2006) Predator recognition and social facilitation of predator avoidance in coral reef fish Dascyllus marginatus juveniles. Mar Ecol Prog Ser 319: 215-223. |
| [68] | Siebeck UE, Litherland L, Wallis GM (2009) Shape learning and discrimination in reef fish. J Exp Biol 212: 2113-2119. |
| [69] |
Newport C, Wallis G, Reshitnyk Y, et al. (2016) Discrimination of human faces by archerfish (Toxotes catareus). Sci Rep 6: e27523. doi: 10.1038/srep27523
|
| [70] |
Greschner M, Field GD, Li PH, et al. (2014) A polyaxonal amacrine cell population in the primate retina. J Neurosci 34: 3597-3606. doi: 10.1523/JNEUROSCI.3359-13.2014
|
| [71] |
Greschner M, Heitman AK, Field GD, et al. (2016) Identification of a retinal circuit for recurrent suppression using indirect electrical imaging. Curr Biol 26: 1935-1942. doi: 10.1016/j.cub.2016.05.051
|
| [72] | Greene E (2007) Retinal encoding of ultrabrief shape recognition cues. PLoS One 2: e871. |
| [73] | Greene E (2016) Retinal encoding of shape boundaries. JSM Anat Physiol 1: 1002. |
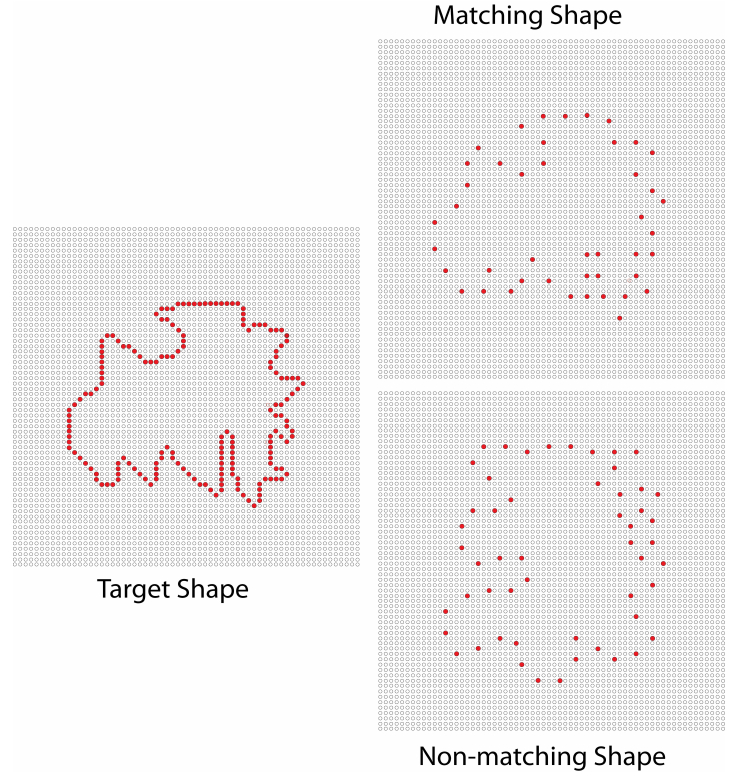
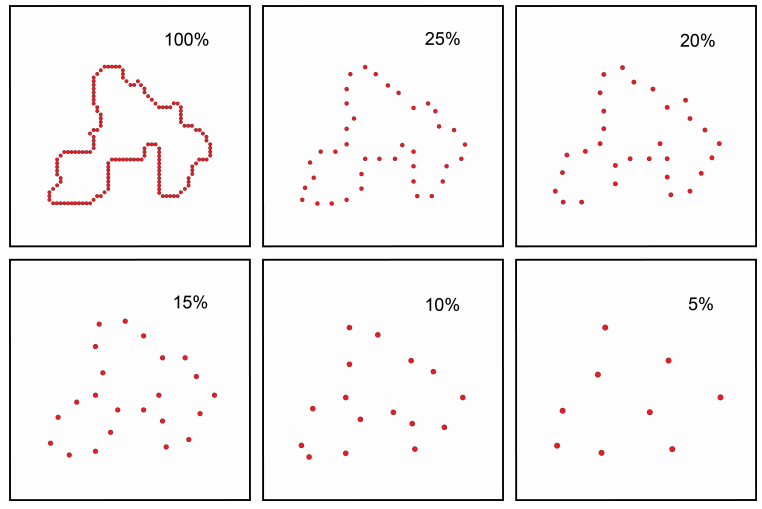
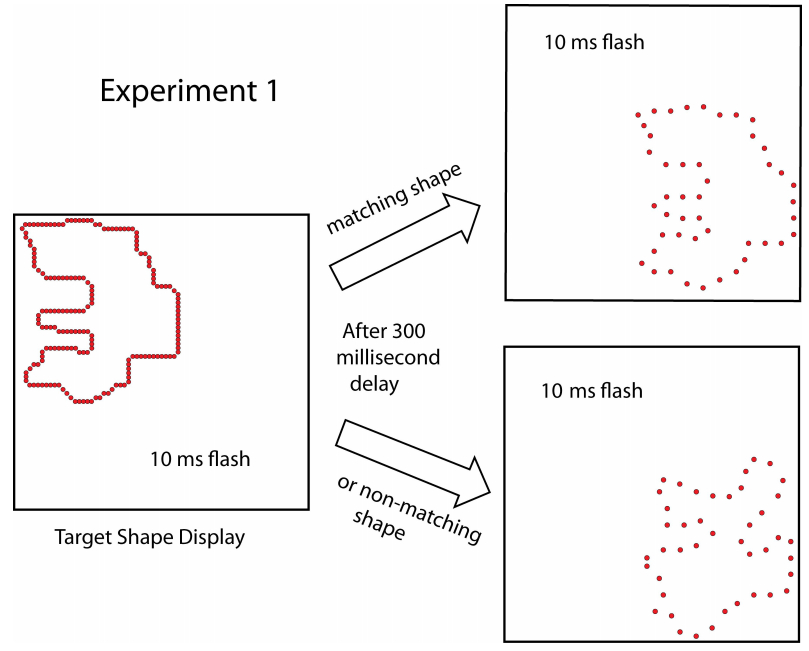
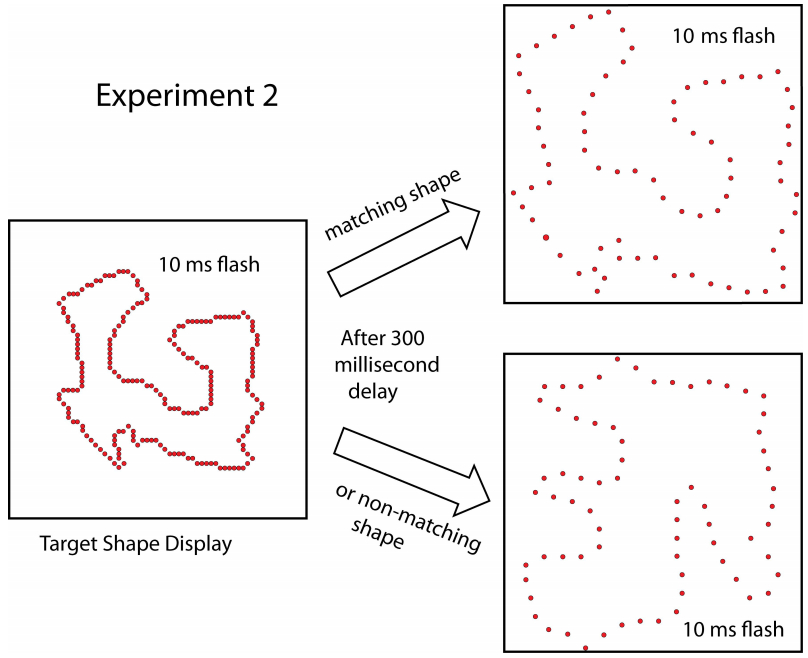
Figures(8)

Ernest Greene, Michael J. Hautus. Demonstrating Invariant Encoding of Shapes Using A Matching Judgment Protocol[J]. AIMS Neuroscience, 2017, 4(3): 120-147. doi: 10.3934/Neuroscience.2017.3.120







 DownLoad:
DownLoad: