Citation: Siri-Maria Kamp, Melissa Lehman, Kenneth J. Malmberg, Emanuel Donchin. A Buffer Model Account of Behavioral and ERP Patterns in the Von Restorff Paradigm[J]. AIMS Neuroscience, 2016, 3(2): 181-202. doi: 10.3934/Neuroscience.2016.2.181

| [1] | Lehman M, Malmberg KJ (2013) A buffer model of memory encoding and temporal correlations in retrieval. Psychol Rev 120: 155-189. doi:10.1037/a0030851 |
| [2] | Lehman M, Malmberg KJ (2009) A global theory of remembering and forgetting from multiple lists. J Exp Psychol Learn Mem Cogn 35: 970-988. doi:10.1037/a0015728 |
| [3] | Atkinson RC, Shiffrin RM (1968) Human memory: A proposed system and its control processes. Psychology Learn Motivation 2: 89-195. |
| [4] | Von Restorff H (1933) Über die Wirkung von Bereichsbildungen im Spurenfeld. Psychologische Forschung 18: 299-342. doi:10.1007/BF02409636 |
| [5] |
Malmberg KJ, Shiffrin RM (2005) The "one-shot" hypothesis for context storage. J Experiment Psychology-Learn Memory Cognition 31: 322-336. doi:Doi 10.1037/0278-7393.31.2.322 doi: 10.1037/0278-7393.31.2.322

|
| [6] |
Schmidt SR (1991) Can we have a distinctive theory of memory? Memory Cognition 19: 523-542. doi: 10.3758/BF03197149
|
| [7] | Sokolov EN (1963) Higher nervous functions; the orienting reflex. Annu Rev Physiol 25: 545-580. doi:10.1146/annurev.ph.25.030163.002553 |
| [8] |
Hunt RR, Lamb CA (2001) What causes the isolation effect?. J Experiment Psychol Learn Memory Cognition 27: 1359. doi: 10.1037/0278-7393.27.6.1359
|
| [9] | Sutton S, Braren M, Zubin J, et al. (1965) Evoked-potential correlates of stimulus uncertainty. Science 150: 1187-1188. doi:10.1126/science.150.3700.1187 |
| [10] | Donchin E (1981) Presidential address, 1980. Surprise!...Surprise? Psychophysiology 18: 493-513. doi:10.1111/j.1469-8986.1981.tb01815.x |
| [11] | Donchin E, Coles MGH (1988) Is the P300 component a manifestation of context updating? Behavior Brain Sci 11: 357. doi:10.1017/s0140525x00058027 |
| [12] | Karis D, Fabiani M, Donchin E (1984) “P300” and memory: Individual differences in the von Restorff effect. Cognitive Psychology 16: 177-216. doi:10.1016/0010-0285(84)90007-0 |
| [13] | Wiswede D, Rüsseler J, Münte TF (2007) Serial position effects in free memory recall--An ERP-study. Biol Psychol 75: 185-193. doi:10.1016/j.biopsycho.2007.02.002 |
| [14] | Duncan-Johnson CC, Donchin E (1977) On quantifying surprise: the variation of event-related potentials with subjective probability. Psychophysiology 14: 456-467. doi:10.1111/j.1469-8986.1977.tb01312.x |
| [15] | Wickens C, Kramer A, Vanasse L, et al. (1983) Performance of concurrent tasks: a psychophysiological analysis of the reciprocity of information-processing resources. Science 221: 1080-1082. doi:10.1126/science.6879207 |
| [16] | Löw A, Rockstroh B, Cohen R, et al. (1999) Determining working memory from ERP topography. Brain Topogr 12: 39-47. doi:10.1023/A:1022229623355 |
| [17] | Ruchkin DS, Canoune HL, Johnson R, et al. (1995) Working memory and preparation elicit different patterns of slow wave event-related brain potentials. Psychophysiology 32: 399-410. doi:10.1111/j.1469-8986.1995.tb01223.x |
| [18] | Mecklinger A, Pfeifer E (1996) Event-related potentials reveal topographical and temporal distinct neuronal activation patterns for spatial and object working memory. Brain Res Cogn Brain Res 4: 211-224. doi:10.1016/S0926-6410(96)00034-1 |
| [19] | Johnson R (1995) Event-related potential insights into the neurobiology of memory systems. In F. Boller & J. Grafman (Eds.), Handbook of neuropsychology (Vol. 10, pp. 135-163). Amsterdam: Elsevier. |
| [20] | Münte TF, Schiltz K, Kutas M (1998) When temporal terms belie conceptual order. Nature 395: 71-73. doi:10.1038/25731 |
| [21] | Lehnert G, Zimmer HD (2008) Common coding of auditory and visual spatial information in working memory. Brain Res 1230: 158-167. doi:10.1016/j.brainres.2008.07.005 |
| [22] | Jäger T, Mecklinger A, Kipp KH (2006) Intra- and inter-item associations doubly dissociate the electrophysiological correlates of familiarity and recollection. Neuron 52: 535-545. doi:10.1016/j.neuron.2006.09.013 |
| [23] | Kamp S.-M, Zimmer HD (2015) Contributions of attention and elaboration to associative encoding in young and older adults. Neuropsychologia 75: 252-264. doi:10.1016/j.neuropsychologia.2015.06.026 |
| [24] | Kim ASN, Vallesi A, Picton TW, et al. (2009) Cognitive association formation in episodic memory: evidence from event-related potentials. Neuropsychologia 47: 3162-3173. doi:10.1016/j.neuropsychologia.2009.07.015 |
| [25] | Fabiani M, Donchin E (1995) Encoding processes and memory organization: a model of the von Restorff effect. J Exp Psychol Learn Mem Cogn 21: 224-240. doi:10.1037/0278-7393.21.1.224 |
| [26] | Fabiani M, Karis D, Donchin E (1990) Effects of mnemonic strategy manipulation in a Von Restorff paradigm. Electroencephalogr Clin Neurophysiol 75: 22-35. doi:10.1016/0013-4694(90)90149-E |
| [27] | Raaijmakers JG, Shiffrin RM (1981) Search of associative memory. Psychological Rev 88: 93. |
| [28] | Kahana MJ (1996) Associative retrieval processes in free recall. Mem Cognit 24: 103-109. doi:10.3758/BF03197276 |
| [29] | Luck SJ, Vogel EK (1997) The capacity of visual working memory for features and conjunctions. Nature 390: 279-281. doi:10.1038/36846 |
| [30] | Kamp S.-M, Brumback T, Donchin E (2013) The component structure of ERP subsequent memory effects in the Von Restorff paradigm and the word frequency effect in recall. Psychophysiology 50: 1079-1093. doi:10.1111/psyp.12090 |
| [31] | Kamp S.-M, Potts GF, Donchin E (2015) On the role of distinctiveness and semantic expectancies in episodic encoding of emotional words. Psychophysiology 52: 1599-1609. |
| [32] | Francis WN, Kucera H (1982) Frequency Analysis of English Usage: Lexicon and Grammar: Boston: Houghton Mifflin. |
| [33] | Dien J (2010) The ERP PCA Toolkit: an open source program for advanced statistical analysis of event-related potential data. J Neurosci Methods 187: 138-145. doi:10.1016/j.jneumeth.2009.12.009 |
| [34] | Spencer KM, Dien J, Donchin E (1999) A componential analysis of the ERP elicited by novel events using a dense electrode array. Psychophysiology 36: 409-414. doi:10.1017/S0048577299981180 |
| [35] | Kamp S.-M, Donchin E (2015) ERP and pupil responses to deviance in an oddball paradigm. Psychophysiology 52: 460-471. doi:10.1111/psyp.12378 |
| [36] | Wiswede D, Rüsseler J, Hasselbach S, et al. (2006) Memory recall in arousing situation—an emotional von Restorff effect? BMC Neurosci 7: 57. doi:10.1186/1471-2202-7-57 |
| [37] | Kutas M, Federmeier KD (2011) Thirty years and counting: finding meaning in the N400 component of the event-related brain potential (ERP). Annu Rev Psychol 62: 621-647. doi:10.1146/annurev.psych.093008.131123 |
| [38] | Dunlosky J, Hunt RR, Clark E (2000) Is perceptual salience needed in explanations of the Von Restorff effect? J Experiment Psychol Learn Memory Cognition 26: 649-657. |
| [39] | Nairne JS (2006) Modeling Distinctiveness: Implications for general memory theory. R.R. Hunt & J. B. Worthen (Eds.). New York: Oxford University Press. |
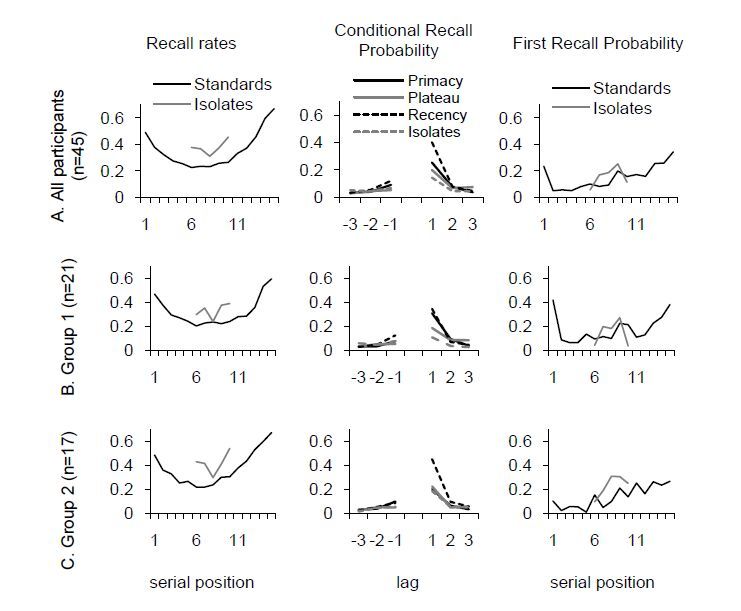
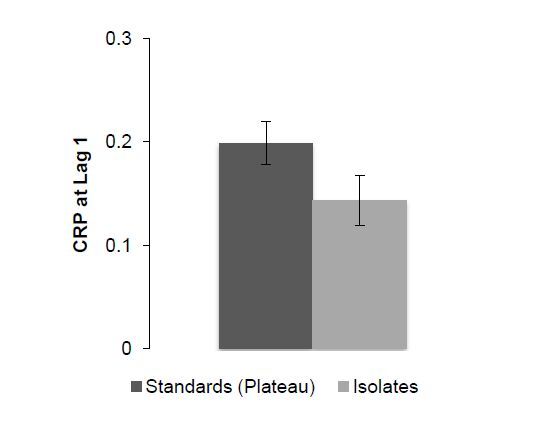
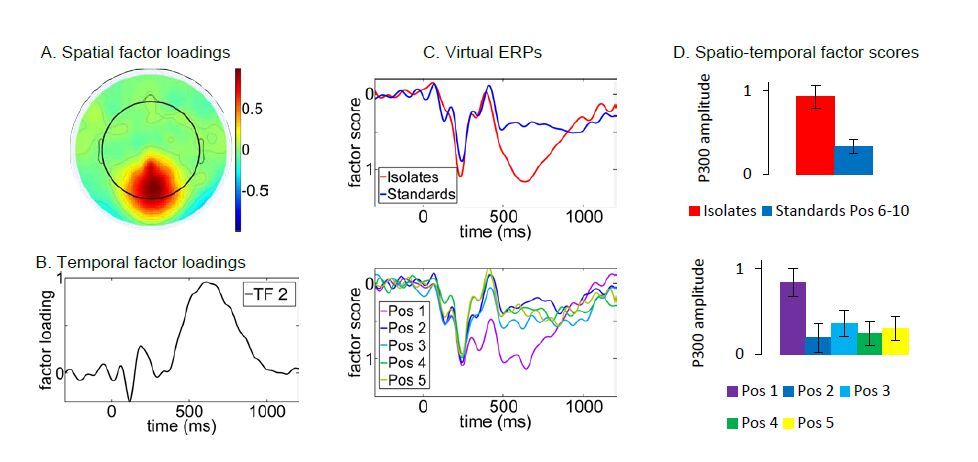
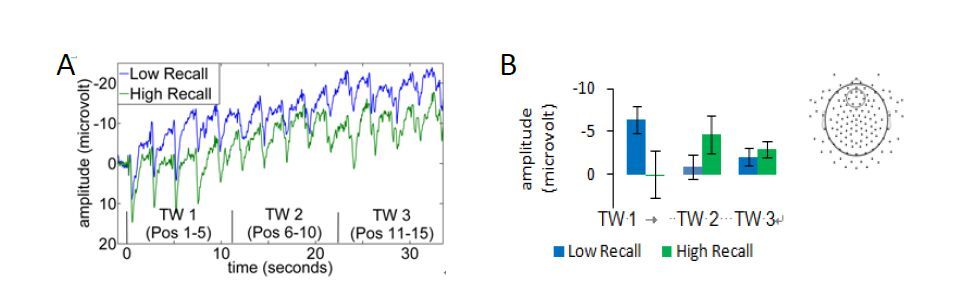
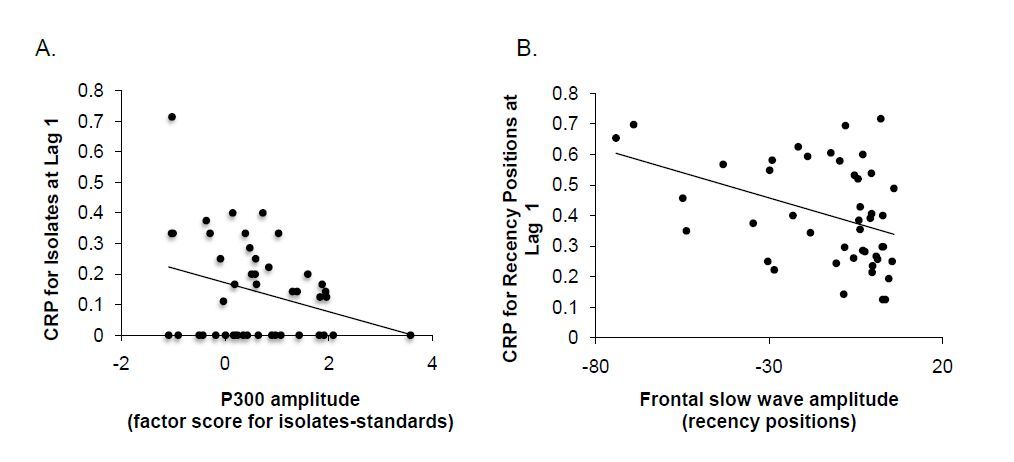
Figures(5)

Siri-Maria Kamp, Melissa Lehman, Kenneth J. Malmberg, Emanuel Donchin. A Buffer Model Account of Behavioral and ERP Patterns in the Von Restorff Paradigm[J]. AIMS Neuroscience, 2016, 3(2): 181-202. doi: 10.3934/Neuroscience.2016.2.181







 DownLoad:
DownLoad: