Human communication and language skills rely heavily on the ability to detect and process auditory inputs. This paper reviews possible applications of the event-related potential (ERP) technique to the study of cortical mechanisms supporting human auditory processing, including speech stimuli. Following a brief introduction to the ERP methodology, the remaining sections focus on demonstrating how ERPs can be used in humans to address research questions related to cortical organization, maturation and plasticity, as well as the effects of sensory deprivation, and multisensory interactions. The review is intended to serve as a primer for researchers interested in using ERPs for the study of the human auditory system.
Citation: Alexandra P. Key. Human Auditory Processing: Insights from Cortical Event-related Potentials[J]. AIMS Neuroscience, 2016, 3(2): 141-162. doi: 10.3934/Neuroscience.2016.2.141
Human communication and language skills rely heavily on the ability to detect and process auditory inputs. This paper reviews possible applications of the event-related potential (ERP) technique to the study of cortical mechanisms supporting human auditory processing, including speech stimuli. Following a brief introduction to the ERP methodology, the remaining sections focus on demonstrating how ERPs can be used in humans to address research questions related to cortical organization, maturation and plasticity, as well as the effects of sensory deprivation, and multisensory interactions. The review is intended to serve as a primer for researchers interested in using ERPs for the study of the human auditory system.

| [1] |
Skoe E, Kraus N. (2010) Auditory brain stem response to complex sounds: A tutorial. Ear Hear 31: 302-324. doi: 10.1097/AUD.0b013e3181cdb272

|
| [2] | Bidelman GM, Moreno S, Alain C (2013) Tracing the emergence of categorical speech perception in the human auditory system. Hum Brain Map J 79: 201-212. |
| [3] | Rockstroh B, Elbert T, Birbaumer N, et al. (1982) Slow brain potentials and behavior. Baltimore, MD: Urban-Schwarzenberg; 1982. |
| [4] |
Giraud A-L, Poeppel D (2012) Cortical oscillations and speech processing: emerging computational principles and operations. Nat Neurosci 15: 511-517. doi: 10.1038/nn.3063
|
| [5] | Jochaut D, Lehongre K, Saitovitch A, et al. (2015) Atypical coordination of cortical oscillations in response to speech in autism. Front Hum Neurosci 27: 9. |
| [6] | Rüsseler J, Nager W, Möbes J, et al. (2005) Cognitive adaptations and neuroplasticity: Lessons from event-related brain potentials. In: Heil P, Schleich H, Budinger E, Konig R, editors. The auditory cortex: A synthesis of human and animal research. Mahwah, NJ: Lawrence Erlbaum Associates, Inc; 2005. pp. 467-485. |
| [7] | Zani A, Proverbio AM (2003) Cognitive electrophysiology of mind and brain. In: Zani A, Proverbio AM, editors. The Cognitive Electrophysiology of Mind and Brain. San Diego, CA: Academic Press; 2003. pp. 3–36. |
| [8] |
Yoder PJ, Camarata S, Camarata M, et al. (2006) Association between differentiated processing of syllables and comprehension of grammatical morphology in children with Down syndrome. Am J Ment Retard 111: 138-152. doi: 10.1352/0895-8017(2006)111[138:ABDPOS]2.0.CO;2
|
| [9] | Luck SJ (2005) An Introduction to the Event-Related Potential Technique. The MIT Press. |
| [10] | Luck SJ (2014) An Introduction to the event-related potential technique. 2nd ed. The MIT Press. |
| [11] |
Eggermont JJ, Ponton CW (2002) The neurophysiology of auditory perception: from single units to evoked potentials. Audiol Neurootol 7: 71-99. doi: 10.1159/000057656
|
| [12] | Fishman YI (2013) The Mechanisms and Meaning of the Mismatch Negativity. Brain Topogr 27: 500-526. |
| [13] |
Tomé D, Barbosa F, Nowak K, et al. (2015) The development of the N1 and N2 components in auditory oddball paradigms: a systematic review with narrative analysis and suggested normative values. J Neural Transmiss 122: 375-391. doi: 10.1007/s00702-014-1258-3
|
| [14] |
Key APF, Dove GO, Maguire MJ (2005) Linking Brainwaves to the Brain: An ERP Primer. Develop Neuropsychol 27: 183-215. doi: 10.1207/s15326942dn2702_1
|
| [15] |
Granier-Deferre C, Lecanuet JP, Cohen H, et al. (1985) Feasibility of Prenatal Hearing Test. Acta Oto-Laryngologica 99: 93-101. doi: 10.3109/00016488509121762
|
| [16] |
Moore JK, Guan YL (2001) Cytoarchitectural and axonal maturation in human auditory cortex. J Assoc Res Otolaryngol 2: 297-311. doi: 10.1007/s101620010052
|
| [17] | Ponton C, Eggermont JJ, Khosla D, et al. (2013) Maturation of human central auditory system activity: separating auditory evoked potentials by dipole source modeling. Clin Neurophysiol 113: 407-420. |
| [18] |
Moore JK, Niparko JK, Perazzo LM, et al. (1997) Effect of adult-onset deafness on the human central auditory system. Ann Otol Rhinol Laryngol 106: 385-390. doi: 10.1177/000348949710600505
|
| [19] |
Ponton CW, Eggermont JJ, Kwong B, et al. (2000) Maturation of human central auditory system activity: evidence from multi-channel evoked potentials. Clin Neurophysiol 111: 220-236. doi: 10.1016/S1388-2457(99)00236-9
|
| [20] | Rotteveel JJ, de Graaf R, Stegeman DF, et al. (1987) The maturation of the central auditory conduction in preterm infants until three months post term. V. The auditory cortical response (ACR). Hear Res 27: 95-110. |
| [21] |
Holst M, Eswaran H, Lowery C, et al. (2005) Development of auditory evoked fields in human fetuses and newborns: A longitudinal MEG study. Clin Neurophysiol 116: 1949-1955. doi: 10.1016/j.clinph.2005.04.008
|
| [22] |
Moller AR, Rollins PR (2002) The non-classical auditory pathways are involved in hearing in children but not in adults. Neurosci Letter 319: 41-44. doi: 10.1016/S0304-3940(01)02516-2
|
| [23] |
Gordon KA, Tanaka S, Wong DDE, et al. (2008) Characterizing responses from auditory cortex in young people with several years of cochlear implant experience. Clin Neurophysiol 119: 2347-2362. doi: 10.1016/j.clinph.2008.06.013
|
| [24] |
Kushnerenko E, Čeponienė R, Balan P, et al. (2002) Maturation of the auditory change detection response in infants: a longitudinal ERP study. NeuroReport 13: 1843–1848. doi: 10.1097/00001756-200210280-00002
|
| [25] |
Trainor L, McFadden M, Hodgson L, et al. (2003) Changes in auditory cortex and the development of mismatch negativity between 2 and 6 months of age. Int J Psychophysiol 51: 5-15. doi: 10.1016/S0167-8760(03)00148-X
|
| [26] |
Wunderlich JL, Cone-Wesson BK (2006) Maturation of CAEP in infants and children: A review. Hear Res 212: 212-223. doi: 10.1016/j.heares.2005.11.008
|
| [27] | Molfese DL, Molfese VJ (2000) The continuum of language development during infancy and early childhood: Electrophysiological correlates. In: Rovee-Collier C, Lipsitt L, Hayne H, editors. Progress in infancy research. Lawrence Erlbaum Associates. pp. 251-287. |
| [28] | Nanova P, Lyamova L, Hadjigeorgieva M, et al. (2008) Gender-specific development of auditory information processing in children: An ERP study. Clin Neurophysiol 119: 1992-2003. |
| [29] | Key APF, Lambert EW, Aschner JL, et al. (2012) Influence of gestational age and postnatal age on speech sound processing in NICU infants. Psychophysiology 49: 720-731. |
| [30] |
Pang EW, Taylor MJ (2000) Tracking the development of the N1 from age 3 to adulthood: an examination of speech and non-speech stimuli. Clin Neurophysiol 111: 388-397. doi: 10.1016/S1388-2457(99)00259-X
|
| [31] |
Ponton CW, Don M, Eggermont JJ, et al. (1996) Auditory system plasticity in children after long periods of complete deafness. NeuroReport 8: 61. doi: 10.1097/00001756-199612200-00013
|
| [32] |
Ponton CW, Don M, Eggermont JJ, et al. (1996) Maturation of human cortical auditory function: differences between normal-hearing children and children with cochlear implants. Ear Hear 17: 430-437. doi: 10.1097/00003446-199610000-00009
|
| [33] |
Paetau R, Ahonen A, Salonen O, et al. (1995) Auditory evoked magnetic fields to tones and pseudowords in healthy children and adults. J Clin Neurophysiol 12: 177-185. doi: 10.1097/00004691-199503000-00008
|
| [34] |
Sharma A, Kraus N, McGee TJ, et al. (1997) Developmental changes in P1 and N1 central auditory responses elicited by consonant-vowel syllables. Electroencephalography clin Neurophysiol 104: 540-545. doi: 10.1016/S0168-5597(97)00050-6
|
| [35] |
Gilley PM, Sharma A, Dorman M, et al. (2005) Developmental changes in refractoriness of the cortical auditory evoked potential. Clin Neurophysiol 116: 648-657. doi: 10.1016/j.clinph.2004.09.009
|
| [36] |
Kaas JH, Hackett TA, Tramo MJ, et al. (1999) Auditory processing in primate cerebral cortex. Current Opinion Neurobiol 9: 164-170. doi: 10.1016/S0959-4388(99)80022-1
|
| [37] |
Morel A, Garraghty PE, Kaas JH (1993) Tonotopic organization, architectonic fields, and connections of auditory cortex in macaque monkeys. J Comp Neurol 335: 437-459. doi: 10.1002/cne.903350312
|
| [38] | Recanzone GH, Guard DC, Phan ML (2000) Frequency and intensity response properties of single neurons in the auditory cortex of the behaving macaque monkey. J Neurophysiol 83: 2315-2331. |
| [39] |
Rapin I, Schimmel H, Tourk LM, et al. (1966) Evoked responses to clicks and tones of varying intensity in waking adults. Electroencephalography Clin Neurophysiol 21: 335-344. doi: 10.1016/0013-4694(66)90039-3
|
| [40] |
Beagley HA, Knight JJ (1967) Changes in Auditory Evoked Response with Intensity. J Laryngology Otology 81: 861-873. doi: 10.1017/S0022215100067815
|
| [41] |
Madell JR, Goldstein R (1972) Relation between loudness and the amplitude of the early components of the averaged electroencephalic response. J Speech Hear Res 15: 134-141. doi: 10.1044/jshr.1501.134
|
| [42] |
Picton TW, Hillyard SA, Krausz HI, et al. (1974) Human auditory evoked potentials. I. Evaluation of components. Electroencephalography Clin Neurophysiol 36: 179-190. doi: 10.1016/0013-4694(74)90155-2
|
| [43] |
Billings CJ, Tremblay KL, Souza PE, et al. (2007) Effects of hearing aid amplification and stimulus intensity on cortical auditory evoked potentials. Audiol Neurotol 12: 234-246. doi: 10.1159/000101331
|
| [44] |
Garinis AC, Cone-Wesson BK (2007) Effects of stimulus level on cortical auditory event-related potentials evoked by speech. J Am Aca Audiology 18: 107-116. doi: 10.3766/jaaa.18.2.3
|
| [45] | Aitkin LM, Merzenich MM, Irvine DR, et al. (2013) Frequency representation in auditory cortex of the common marmoset (Callithrix jacchus jacchus). J Comp Neurol 252: 175-185. |
| [46] |
Picton TW, Woods DL, Proulx GB (1978) Human auditory sustained potentials. II. Stimulus relationships. Electroencephalography Clin Neurophysiol 45: 198-210. doi: 10.1016/0013-4694(78)90004-4
|
| [47] |
Crottaz-Herbette S, Ragot R (2000) Perception of complex sounds: N1 latency codes pitch and topography codes spectra. Clin Neurophysiol 111: 1759-1766. doi: 10.1016/S1388-2457(00)00422-3
|
| [48] |
Verkindt C, Bertrand O, Perrin F, et al. (1995) Tonotopic organization of the human auditory cortex: N100 topography and multiple dipole model analysis. Electroencephalography Clin Neurophysiol 96:143–56. doi: 10.1016/0168-5597(94)00242-7
|
| [49] |
Wunderlich JL, Cone-Wesson BK (2001) Effects of stimulus frequency and complexity on the mismatch negativity and other components of the cortical auditory-evoked potential. J Acoust Soc Am 109: 1526-1537. doi: 10.1121/1.1349184
|
| [50] | Bertrand O, Perrin F, Pernier J (1991) Evidence for a tonotopic organization of the auditory cortex observed with auditory evoked potentials. Acta Oto-Laryngologica 111: 116-123. |
| [51] |
Pantev C, Bertrand O, Eulitz C, et al. (1995) Specific tonotopic organizations of different areas of the human auditory cortex revealed by simultaneous magnetic and electric recordings. Electroencephalography Clin Neurophysiol 94: 26-40. doi: 10.1016/0013-4694(94)00209-4
|
| [52] |
Lütkenhöner B, Steinsträter O (1998) High-precision neuromagnetic study of the functional organization of the human auditory cortex. Audiol Neurootol 3: 191-213. doi: 10.1159/000013790
|
| [53] |
Shestakova A, Brattico E, Soloviev A, et al. (2004) Orderly cortical representation of vowel categories presented by multiple exemplars. Brain Res Cogn Brain Res 21: 342-350. doi: 10.1016/j.cogbrainres.2004.06.011
|
| [54] |
Näätänen R, Paavilainen P, Rinne T, et al. (2007) The mismatch negativity (MMN) in basic research of central auditory processing: A review. Clin Neurophysiol 118: 2544-2590. doi: 10.1016/j.clinph.2007.04.026
|
| [55] | May PJC, Tiitinen H (2010) Mismatch negativity (MMN), the deviance-elicited auditory deflection, explained. Psychophysiology 47: 66-122. |
| [56] |
Garrido MI, Kilner JM, Stephan KE, et al. (2009) The mismatch negativity: A review of underlying mechanisms. Clin Neurophysiol 120: 453-463. doi: 10.1016/j.clinph.2008.11.029
|
| [57] |
Cheour-Luhtanen M, Alho K, Sainio K, et al. (1996) The ontogenetically earliest discriminative response of the human brain. Psychophysiology 33: 478-481. doi: 10.1111/j.1469-8986.1996.tb01074.x
|
| [58] | Leppänen PHT, Richardson U, Pihko E, et al. (2002) Brain responses to changes in speech sound durations differ between infants with and without familial risk for dyslexia. Develop Neuropsychol 22: 407-422. |
| [59] |
Alho K, Sainio K, Sajaniemi N, et al. (1990) Event-related brain potential of human newborns to pitch change of an acoustic stimulus. Electroencephalography Clin Neurophysiol 77: 151-155. doi: 10.1016/0168-5597(90)90031-8
|
| [60] |
Čeponienė R, Kushnerenko E, Fellman V, et al. (2002) Event-related potential features indexing central auditory discrimination by newborns. Brain Res Cogn Brain Res 13: 101-113. doi: 10.1016/S0926-6410(01)00093-3
|
| [61] | Cheour M, Kushnerenko E, Čeponienė R, et al. (2002) Electric brain responses obtained from newborn infants to changes in duration in complex harmonic tones. Develop Neuropsychol 22: 471-479. |
| [62] |
Kushnerenko E, Čeponienė R, Fellman V, et al. (2001) Event-related potential correlates of sound duration: similar pattern from birth to adulthood. NeuroReport 12: 3777-3781. doi: 10.1097/00001756-200112040-00035
|
| [63] |
Martynova O, Kirjavainen J, Cheour M (2003) Mismatch negativity and late discriminative negativity in sleeping human newborns. Neurosci Letter 340: 75-78. doi: 10.1016/S0304-3940(02)01401-5
|
| [64] |
Brannon EM, Roussel LW, Meck WH, et al. (2004) Timing in the baby brain. Brain Res Cogn Brain Res 21: 227-233. doi: 10.1016/j.cogbrainres.2004.04.007
|
| [65] |
Friederici AD, Friedrich M, Weber C (2002) Neural manifestation of cognitive and precognitive mismatch detection in early infancy. NeuroReport 13: 1251-1254. doi: 10.1097/00001756-200207190-00006
|
| [66] | Pihko E, Leppänen PH, Eklund KM, et al. (1999) Cortical responses of infants with and without a genetic risk for dyslexia: I. Age effects. NeuroReport 10: 901-905. |
| [67] | Steinschneider M, Volkov IO, Noh MD, et al. (1999) Temporal encoding of the voice onset time phonetic parameter by field potentials recorded directly from human auditory cortex. J Neurophysiol 82: 2346-2357. |
| [68] |
Simos PG, Molfese DL, Brenden RA (1997) Behavioral and Electrophysiological Indices of Voicing-Cue Discrimination: Laterality Patterns and Development. Brain Language 57: 122-150. doi: 10.1006/brln.1997.1836
|
| [69] | Molfese DL, Papanicolaou A, Hess TM, et al. (1979) Neuroelectrical correlates of semantic processes. Evoked brain potentials and behavior. Plenum Pub Cor: 89-106. |
| [70] | Molfese DL, Molfese VJ (1979) Hemisphere and stimulus differences as reflected in the cortical responses of newborn infants to speech stimuli. Develop Psychol 15: 505-511. |
| [71] |
Segalowitz SJ, Cohen H (1989) Right hemisphere EEG sensitivity to speech. Brain Language 37: 220-231. doi: 10.1016/0093-934X(89)90016-3
|
| [72] | Dehaene-Lambertz G, Baillet S (1998) A phonological representation in the infant brain. NeuroReport 9: 1885. |
| [73] |
Dehaene-Lambertz G, Dehaene S (1994) Speed and cerebral correlates of syllable discrimination in infants. Nature 370: 292-295. doi: 10.1038/370292a0
|
| [74] | Dehaene-Lambertz G (2000) Cerebral specialization for speech and non-speech stimuli in infants. J Cogn Neurosci 12: 449-460. |
| [75] | Molfese DL (2000) Predicting dyslexia at 8 years of age using neonatal brain responses. Brain Language 72: 238-245. |
| [76] |
Kraus N, McGee T, Micco A, et al. (1993) Mismatch negativity in school-age children to speech stimuli that are just perceptibly different. Electroencephalography Clin Neurophysiol 88: 123-130. doi: 10.1016/0168-5597(93)90063-U
|
| [77] | Molfese D, Key A, Maguire M, et al. (2005) Event-related evoked potentials (ERPs) in speech perception. In: Pisoni D, Remez R, editors. The Handbook of Speech Perception. Malden, MA: pp. 99-121. |
| [78] |
Zhang LI, Bao S, Merzenich MM (2002) Disruption of primary auditory cortex by synchronous auditory inputs during a critical period. Proc Natl Acad Sci USA 99: 2309-2314. doi: 10.1073/pnas.261707398
|
| [79] | Stanton SG, Harrison RV (1996) Abnormal cochleotopic organization in the auditory cortex of cats reared in a frequency augmented environment. Aud Neurosci 2: 97-107. |
| [80] |
Nakahara H, Zhang LI, Merzenich MM (2004) Specialization of primary auditory cortex processing by sound exposure in the “critical period.” Proceed Nation Aca Sci 101: 7170-7174. doi: 10.1073/pnas.0401196101
|
| [81] |
deRegnier R-A, Wewerka S, Georgieff MK, et al. (2002) Influences of postconceptional age and postnatal experience on the development of auditory recognition memory in the newborn infant. Dev Psychobiol 41: 216-225. doi: 10.1002/dev.10070
|
| [82] | Bisiacchi PS, Mento G, Suppiej A (2009) Cortical auditory processing in preterm newborns: An ERP study. Biolog Psychol 82: 176-185. |
| [83] |
Fellman V, Kushnerenko E, Mikkola K, et al. (2004) Atypical auditory event-related potentials in preterm infants during the first year of life: A possible sign of cognitive dysfunction? Pediatric Res 56: 291-297. doi: 10.1203/01.PDR.0000132750.97066.B9
|
| [84] | Cheour M, Martynova O, Näätänen R, et al. (2002) Speech sounds learned by sleeping newborns. Nature 415: 599-600. |
| [85] |
Cheour M, Čeponienė R, Lehtokoski A, et al. (1998) Development of language-specific phoneme representations in the infant brain. Nat Neurosci 1: 351-353. doi: 10.1038/1561
|
| [86] |
Rivera-Gaxiola M, Silva Pereyra J, Kuhl PK (2005) Brain potentials to native and non-native speech contrasts in 7-and 11-month-old American infants. Develop Sci 8: 162-172. doi: 10.1111/j.1467-7687.2005.00403.x
|
| [87] | Aaltonen O, Niemi P, Nyrke T, et al. (1987) Event-related brain potentials and the perception of a phonetic continuum. Biolog Psychol 24: 197-207. |
| [88] |
Dehaene-Lambertz G (1997) Electrophysiological correlates of categorical phoneme perception in adults. NeuroReport 8: 919-924. doi: 10.1097/00001756-199703030-00021
|
| [89] |
Rivera-Gaxiola M, Csibra G, Johnson MH (2000) Electrophysiological correlates of cross-linguistic speech perception in native English speakers. Behav Brain Res 111: 13-23. doi: 10.1016/S0166-4328(00)00139-X
|
| [90] |
Sharma A, Dorman MF (2000) Neurophysiologic correlates of cross-language phonetic perception. J Acoust Soc Am 107: 2697. doi: 10.1121/1.428655
|
| [91] |
Cheour M, Shestakova A, Alku P, et al. (2002) Mismatch negativity shows that 3-6-year-old children can learn to discriminate non-native speech sounds within two months. Neurosci Letter 325: 187-190. doi: 10.1016/S0304-3940(02)00269-0
|
| [92] | Tremblay K, Kraus N, McGee T, et al. (2001) Central auditory plasticity: changes in the N1-P2 complex after speech-sound training. Ear Hear 22: 79-90. |
| [93] |
Tremblay KL, Kraus N et al. (2002) Auditory training induces asymmetrical changes in cortical neural activity. J Speech Lang Hear Re 45: 564-572. doi: 10.1044/1092-4388(2002/045)
|
| [94] |
Beitel RE, Schreiner CE, Cheung SW, et al. (2003) Reward-dependent plasticity in the primary auditory cortex of adult monkeys trained to discriminate temporally modulated signals. Proc Natl Acad Sci USA 100: 11070-11075. doi: 10.1073/pnas.1334187100
|
| [95] |
Reinke KS, He Y, Wang C, et al. (2003) Perceptual learning modulates sensory evoked response during vowel segregation. Cogn Brain Res 17: 781-791. doi: 10.1016/S0926-6410(03)00202-7
|
| [96] |
Sheehan KA, McArthur GM, Bishop DVM (2005) Is discrimination training necessary to cause changes in the P2 auditory event-related brain potential to speech sounds? Cogn Brain Res 25: 547-553. doi: 10.1016/j.cogbrainres.2005.08.007
|
| [97] |
Tremblay K, Ross B (2007) Effects of age and age-related hearing loss on the brain. J Commu Disorder 40: 305-312. doi: 10.1016/j.jcomdis.2007.03.008
|
| [98] |
Mendelson JR, Ricketts C (2001) Age-related temporal processing speed deterioration in auditory cortex. Hear Res 158: 84-94. doi: 10.1016/S0378-5955(01)00294-5
|
| [99] |
Papanicolaou AC, Loring DW, Eisenberg HM (1984) Age-related differences in recovery cycle of auditory evoked potentials. Neurobiol Aging 5: 291-295. doi: 10.1016/0197-4580(84)90005-8
|
| [100] | Illing R-B (2004) Maturation and plasticity of the central auditory system. Acta Oto-Laryngologica 124: 6-10. |
| [101] |
Kral A (2004) Postnatal cortical development in congenital auditory deprivation. Cerebral Cortex 15: 552-562. doi: 10.1093/cercor/bhh156
|
| [102] | Sharma A, Tobey E, Dorman M, et al. (2004) Central auditory maturation and babbling development in infants with cochlear implants. Arch Otolaryngol Head Neck Surg 130: 511-516. |
| [103] | Weitzman L, Graziani L, Duhamel L (1967) Maturation and topography of the auditory evoked response of the prematurely born infant. Electroencephalography Clin Neurophysiol 23: 82-83. |
| [104] |
Polen SB (1984) Auditory event related potentials. Seminars Hear 5: 127-141. doi: 10.1055/s-0028-1095227
|
| [105] |
Wall LG, Dalebout SD, Davidson SA, et al. (1991) Effect of Hearing Impairment on Event-Related Potentials for Tone and Speech Distinctions. Folia Phoniatr Logop 43: 265-274. doi: 10.1159/000266137
|
| [106] | Oates PA, Kurtzberg D, Stapells DR (2002) Effects of sensorineural hearing loss on cortical event-related potential and behavioral measures of speech-sound processing. Ear Hear 23:399-415. |
| [107] | Campbell J, Sharma A (2013) Compensatory changes in cortical resource allocation in adults with hearing loss. Front Syst Neurosci 7: 71. |
| [108] |
Bertoli S, Probst R, Bodmer D (2011) Late auditory evoked potentials in elderly long-term hearing-aid users with unilateral or bilateral fittings. Hear Res 280: 58-69. doi: 10.1016/j.heares.2011.04.013
|
| [109] |
Noreña AJ, Eggermont JJ (2005) Enriched acoustic environment after noise trauma reduces hearing loss and prevents cortical map reorganization. J Neurosci 25: 699-705. doi: 10.1523/JNEUROSCI.2226-04.2005
|
| [110] |
Korczak PA, Kurtzberg D, Stapells DR (2005) Effects of sensorineural hearing loss and personal hearing aids on cortical event-related potential and behavioral measures of speech-sound processing. Ear Hear 26: 165-185. doi: 10.1097/00003446-200504000-00005
|
| [111] |
Sharma A, Dorman MF, Spahr AJ (2002) A sensitive period for the development of the central auditory system in children with cochlear implants: Implications for age of implantation. Ear Hear 23: 532-539. doi: 10.1097/00003446-200212000-00004
|
| [112] |
Jordan K, Schmidt A, Plotz K, et al. (1997) Auditory event-related potentials in post- and prelingually deaf cochlear implant recipients. Am J Otology 18: S116-117. doi: 10.1016/S0196-0709(97)90099-5
|
| [113] | Sharma A, Dorman MF, Kral A (2005) The influence of a sensitive period on central auditory development in children with unilateral and bilateral cochlear implants. Hear Res 203: 134-143. |
| [114] |
Sharma A, Dorman MF, Spahr AJ (2002) Rapid development of cortical auditory evoked potentials after early cochlear implantation. NeuroReport 13: 1365-1368. doi: 10.1097/00001756-200207190-00030
|
| [115] |
Dorman MF, Sharma A, Gilley P, et al. (2007) Central auditory development: Evidence from CAEP measurements in children fit with cochlear implants. J Commu Disorders 40: 284-294. doi: 10.1016/j.jcomdis.2007.03.007
|
| [116] |
Ponton CW, Eggermont JJ (2001) Of kittens and kids: altered cortical maturation following profound deafness and cochlear implant use. Audiol Neurootol 6: 363-380. doi: 10.1159/000046846
|
| [117] | Sharma A, Dorman M, Spahr A, et al. (2002) Early cochlear implantation in children allows normal development of central auditory pathways. Ann Otol Rhinol Laryngol Suppl 189: 38-41. |
| [118] |
Johnson JM (2009) Late Auditory Event-Related Potentials in Children With Cochlear Implants: A Review. Develop Neuropsychol 34: 701-720. doi: 10.1080/87565640903265152
|
| [119] |
Finney EM, Fine I, Dobkins KR (2001) Visual stimuli activate auditory cortex in the deaf. Nat Neurosci 4: 1171-1173. doi: 10.1038/nn763
|
| [120] | Lee DS, Lee JS, Oh SH, et al. (2001) Cross-modal plasticity and cochlear implants. Nature 409: 149-150. |
| [121] |
Eggermont JJ, Ponton CW (2003) Auditory-evoked potential studies of cortical maturation in normal hearing and implanted children: Correlations with changes in structure and speech perception. Acta Oto-Laryngologica 123: 249-252. doi: 10.1080/0036554021000028098
|
| [122] | Singh S, Liasis A, Rajput K, et al. (2004) Event-related potentials in pediatric cochlear implant patients. Ear Hear 25: 598-610. |
| [123] |
Martin BA, Kurtzberg D, Stapells DR (1999) The effects of decreased audibility produced by high-pass noise masking on N1 and the mismatch negativity to speech sounds /ba/and/da. J Speech Language Hear Res 42: 271-286. doi: 10.1044/jslhr.4202.271
|
| [124] |
Groenen PAP, Beynon AJ, Snik AFM, et al. (2001) Speech-evoked cortical potentials recognition in cochlear implant users and speech. Scand Audiol 30: 31-40. doi: 10.1080/010503901750069554
|
| [125] | Makhdoum MJ, Groenen PAP, Snik AFM, et al. (1998) Intra-and interindividual correlations between auditory evoked potentials and speech perception in cochlear implant users. Scand Audiol 27: 13-20. |
| [126] | Kelly AS, Purdy SC, Thorne PR (2005) Electrophysiological and speech perception measures of auditory processing in experienced adult cochlear implant users. Clin Neurophysiol 116: 1235-1246. |
| [127] | Sandmann P, Plotz K, Hauthal N, et al. (2015) Rapid bilateral improvement in auditory cortex activity in postlingually deafened adults following cochlear implantation. Clin Neurophysiol 126: 594-607. |
| [128] |
Groenen PA, Makhdoum M, van den Brink JL, et al. (1996) The relation between electric auditory brain stem and cognitive responses and speech perception in cochlear implant users. Acta Oto-Laryngologica 116: 785-790. doi: 10.3109/00016489609137926
|
| [129] | Micco AG, Kraus N, Koch DB, et al. (1995) Speech-evoked cognitive P300 potentials in cochlear implant recipients. Am J Otology 16: 514-520. |
| [130] |
Kraus N, Micco AG, Koch DB, et al. (1993) The mismatch negativity cortical evoked potential elicited by speech in cochlear-implant users. Hear Res 65: 118-124. doi: 10.1016/0378-5955(93)90206-G
|
| [131] |
Bauer PW, Sharma A, Martin K, et al. (2006) Central auditory development in children with bilateral cochlear implants. Arch Otolaryngol Head Neck Surg 132: 1133-1136. doi: 10.1001/archotol.132.10.1133
|
| [132] |
Wallace MT, Ramachandran R, Stein BE (2004) A revised view of sensory cortical parcellation. Proc Natl Acad Sci USA 101: 2167-2172. doi: 10.1073/pnas.0305697101
|
| [133] | Murray MM, Spierer L (2009) Auditory spatio-temporal brain dynamics and their consequences for multisensory interactions in humans. Hear Res 258: 121-133. |
| [134] | Musacchia G, Schroeder CE (2009) Neuronal mechanisms, response dynamics and perceptual functions of multisensory interactions in auditory cortex. Hear Res 258: 72-79. |
| [135] |
Sams M, Aulanko R, Hämäläinen M, et al. (1991) Seeing speech: visual information from lip movements modifies activity in the human auditory cortex. Neurosci Letters 127: 141-145. doi: 10.1016/0304-3940(91)90914-F
|
| [136] |
McGurk H, MacDonald J (1976) Hearing lips and seeing voices. Nature 264: 746-748. doi: 10.1038/264746a0
|
| [137] |
Colin C, Radeau M, Soquet A, et al. (2002) Mismatch negativity evoked by the McGurk-MacDonald effect: a phonetic representation within short-term memory. Clin Neurophysiol 113: 495-506. doi: 10.1016/S1388-2457(02)00024-X
|
| [138] |
Möttönen R, Krause CM, Tiippana K, et al. (2002) Processing of changes in visual speech in the human auditory cortex. Brain Res Cogn Brain Res 13: 417-425. doi: 10.1016/S0926-6410(02)00053-8
|
| [139] |
Talsma D, Doty TJ, Woldorff MG (2006) Selective Attention and audiovisual integration: Is attending to both modalities a prerequisite for early integration? Cerebral Cortex 17: 679-690. doi: 10.1093/cercor/bhk016
|
| [140] |
Boutros NN, Belger A (1999) Midlatency evoked potentials attenuation and augmentation reflect different aspects of sensory gating. Biolog Psychiatry 45: 917-922. doi: 10.1016/S0006-3223(98)00253-4
|
| [141] |
Giard M, Peronnet F (1999) Auditory-visual integration during multimodal object recognition in humans: A behavioral and electrophysiological study. J Cogn Neurosci 11: 473-490. doi: 10.1162/089892999563544
|
| [142] | Li Q, Wu J, Touge T (1991) Audiovisual interaction enhances auditory detection in late stage: an event-related potential study. NeuroReport 21: 173-178. |
| [143] |
Van Wassenhove V, Grant KW, Poeppel D (2005) Visual speech speeds up the neural processing of auditory speech. Proc Natl Acad Sci USA 102: 1181-1186. doi: 10.1073/pnas.0408949102
|
| [144] | Besle J, Fort A, Delpuech C, et al. (2004) Bimodal speech: early suppressive visual effects in human auditory cortex. Europ J Neurosci 20: 2225-2234. |
| [145] |
Besle J, Fischer C, Bidet-Caulet A, et al. (2008) Visual Activation and audiovisual interactions in the auditory cortex during speech perception: Intracranial recordings in humans. J Neurosci 28: 14301-14310. doi: 10.1523/JNEUROSCI.2875-08.2008
|
| [146] | Raij T, Uutela K, Hari R (2000) Audiovisual integration of letters in the human brain. Neuron 28: 617-625. |
| [147] |
Näätänen R, Picton T (1987) The N1 wave of the human electric and magnetic response to sound: a review and an analysis of the component structure. Psychophysiology 24: 375-425. doi: 10.1111/j.1469-8986.1987.tb00311.x
|
| [148] |
Klucharev V (2003) Electrophysiological indicators of phonetic and non-phonetic multisensory interactions during audiovisual speech perception. Cogn Brain Res 18: 65-75. doi: 10.1016/j.cogbrainres.2003.09.004
|
| [149] |
Stekelenburg JJ, Vroomen J (2007) Neural correlates of multisensory integration of ecologically valid audiovisual events. J Cogn Neurosci 19: 1964-1973. doi: 10.1162/jocn.2007.19.12.1964
|
| [150] | Bushara KO, Grafman J, Hallett M (2001) Neural correlates of auditory–visual stimulus onset asynchrony detection. J Neurosci 21: 300-304. |
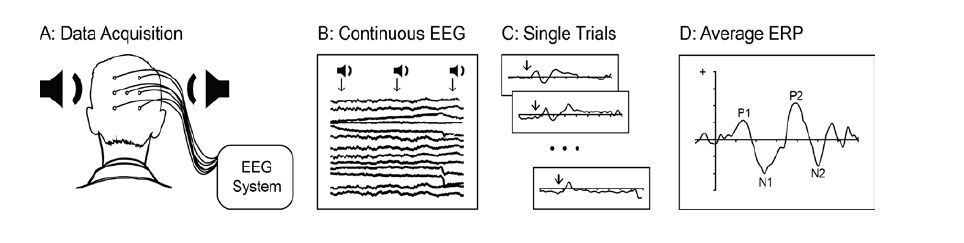
Figures(1)

Alexandra P. Key. Human Auditory Processing: Insights from Cortical Event-related Potentials[J]. AIMS Neuroscience, 2016, 3(2): 141-162. doi: 10.3934/Neuroscience.2016.2.141







 DownLoad:
DownLoad: