Citation: Paul G. Nestor, Toshiyuki Ohtani, James J. Levitt, Dominick T. Newell, Martha E. Shenton, Margaret Niznikiewicz, Robert W. McCarley. Prefrontal Lobe Gray Matter, Cognitive Control and Episodic Memory in Healthy Cognition[J]. AIMS Neuroscience, 2016, 3(3): 338-355. doi: 10.3934/Neuroscience.2016.3.338

| [1] | James W (1890) Attention. The Principles of Psychology I: 427. 10.1037/10538-011 |
| [2] |
Gabrieli JDE (1998) Cognitive neuroscience of human memory. An Rev Psychol 49: 87-115. doi: 10.1146/annurev.psych.49.1.87

|
| [3] |
Kapur S, Craik FIM, Tulving E, et al. (1994) Neuroanatomical correlates of encoding in episodic memory: levels of processing effect. Proc Natl Acad Sci 91: 2008-2011. doi: 10.1073/pnas.91.6.2008
|
| [4] |
Posner MI, Petersen SE, Fox PT, et al. (1988) Localization of cognitive operations in the human brain. Science 240: 1627-1631. DOI: 10.1126/science.3289116 doi: 10.1126/science.3289116
|
| [5] | Posner MI, Petersen SE (1990) The attention system of the human brain. An Rev Neurosci 13: 25-42. |
| [6] |
Petersen SE, Posner MI (2012) The Attention system of the human brain: 20 years after. An Rev Neurosci 35: 73-89. doi:10.1146/annurev-neuro-062111-150525. doi: 10.1146/annurev-neuro-062111-150525
|
| [7] |
Squire LR (1986) Mechanisms of memory. Science 232: 1612-1619. doi:10.1126/science.3086978 doi: 10.1126/science.3086978
|
| [8] |
Squire LR (2004) Memory systems of the brain: A brief history and current perspective. Neurobio Learning Memory 82: 171-177. doi: 10.1016/j.nlm.2004.06.005
|
| [9] |
Squire LR, Zola-Morgan S (1991) The medial temporal love memory system. Science 253: 1380-1386. doi:10.1126/science.1896849. doi: 10.1126/science.1896849
|
| [10] |
Chun MM, Turk-Browne NB (2007) Interactions between attention and memory. Current Opinion Neurobio 17: 177-184. doi: 10.1016/j.conb.2007.03.005. doi: 10.1016/j.conb.2007.03.005
|
| [11] |
Badre D, Poldrack RA, Pare-Blagoev EJ, et al. (2005) Dissociable controlled retrieval and generalized selection mechanisms in ventrolateral prefrontal cortex. Neuron 47: 907-918. doi:10.1016/j.neuron.2005.07.023. doi: 10.1016/j.neuron.2005.07.023
|
| [12] |
Wagner AD, Shannon BJ, Kahn I, et al. (2005) Parietal lobe contributions to episodic memory retrieval. Trends Cogni Sci 9: 445-453. doi:10.1016/j.tics.2005.07.001 doi: 10.1016/j.tics.2005.07.001
|
| [13] |
Cohen JD, Perstein WM, Braver TS, et al. (1997) Temporal dynamics of brain activation during a working memory task. Nature 386: 604-608. doi: 10.1038/386604a0
|
| [14] |
Miller EK, Cohen JD (2001) An integrative theory of prefrontal function. An Rev Neurosci 24: 167-202. doi: 10.1146/annurev.neuro.24.1.167
|
| [15] | Baddeley A (1996) Exploring the central executive. Quarter J Experiment Psychol 49A: 5-28. |
| [16] |
Braver TS (2012) The variable nature of cognitive control: A dual-mechanisms framework. Trends Cogn Sci 16: 106-113. doi: 10.1016/j.tics.2011.12.010
|
| [17] | Elliot R, Dolan RJ, Frith C (2000) Dissociable functions in the medial and lateral orbitofrontal cortex: Evidence from human neuroimaging studies. Cerebral Cortex 10: 308-317. |
| [18] |
Jiang J, Egner T (2014) Using neural pattern classifiers to quantify the modularity of conflict-control mechanisms in the human brain. Cerebral Cortex 24: 1793-1805. doi: 10.1093/cercor/bht029
|
| [19] | Miller EK, Wallis JD (2009) Executive function and higher-order cognition: Definition and neural substrates. In L.R. Squire (ed.) Encyclopedia Neuroscience Vol. 4, pp 99-104. Oxford: Academic Press |
| [20] |
Anderson MC, Spellman BA (1995) On the status of inhibitory mechanisms in cognition: Memory retrieval as a model case. Psychol Rev 102: 68-100. doi:10.1037/0033-295x.102.1.68. doi: 10.1037/0033-295X.102.1.68
|
| [21] |
Bissett PG, Nee DE, Jonides J (2009) Dissociating interference-control processes between memory and response. J Experiment Psychol Learning Memory Cognition 35: 1306-1316. DOI: 10.1037/a0016537. doi: 10.1037/a0016537
|
| [22] |
Nestor PG, Piech R, Allen C, et al. (2005) Retrieval-induced forgetting in schizophrenia. Schizophrenia Res 75: 199-209. doi: 10.1016/j.schres.2005.01.011
|
| [23] |
Depue BE, Curran T, Banich MT (2007) Prefrontal Regions Orchestrate Suppression of Emotional Memories via a Two-Phase Process. Science 317: 215-219. doi:10.1126/science.1139560. doi: 10.1126/science.1139560
|
| [24] |
Peters GJ, David CN, Marcus MD, et al. (2013) The medial prefrontal cortex is critical for memory retrieval and resolving interference. Learning Memory 20: 201-209 doi: 10.1101/lm.029249.112
|
| [25] |
Koechlin E, Hyafil A (2007) Anterior prefrontal function and the limits of human decision making. Science 318: 594-598. doi: 10.1126/science.1142995
|
| [26] | Nestor PG, Ohtani T, Bouix S, et al. (2015) Dissociating prefrontal circuitry in intelligence and memory: Neuropsychological correlates of magnetic resonance and diffusion tensor imaging. Brain Imaging Behavior 9: 839-847. doi: 10.1007/s11682-014-9344-6. |
| [27] |
Ohtani T, Nestor PG, Bouix S, et al. (2014) Medial Frontal White and gray matter contributions to general intelligence. PLoS One 9: e112691. doi:10.137/journal.pone.0112691. doi: 10.1371/journal.pone.0112691
|
| [28] | Nestor PG, Ohtani, Levitt JJ, et al. (2015) Attentional control and intelligence: MRI orbital frontal gray matter and neuropsychological correlates. Behavior Neurology. http://dx.doi.org/10.1155/2015/354186 |
| [29] | Heaton RK (1981) Wisconsin Card Sort Manual. Odessa: Psychological Assessment Resources. |
| [30] | Arbuthnott K, Frank J (2000) Trail Making Test, Part B as a measure of executive control: Validation using a set-switching paradigm. J Clin Experiment Neuropsychol 22: 518-528. |
| [31] | Baddeley AD, Emslie H, Nimmo-Smith I (1994) Doors and People: A test of visual and verbal recall and recognition. Flempton, Bury St. Edmunds: Thames Valley Test Company. |
| [32] | Buschman TJ, Miller EK (2014) Goal-direction and top-down control. Phil Trans R Soc B 369. doi:10.1098/rstb.2013.0471. |
| [33] |
Mansouri FA, Matsumoto K, Tanaka K (2006) Prefrontal cell activities related to monkeys’ success and failure in adapting to rule changes in a Wisconsin Card Sorting Test analog. J Neurosci 26: 2745-2756. doi: 10.1523/JNEUROSCI.5238-05.2006
|
| [34] |
Nestor PG, Kubicki M, Niznikiewicz M, et al. (2008) Neuropsychological disturbance in schizophrenia: A diffusion tensor imaging study. Neuropsychology 22: 246-254. doi: 10.1037/0894-4105.22.2.246
|
| [35] | Manns JR, Squire LR (1999) Impaired recognition memory on the Doors and People Test after damage limited to the hippocampal region. Hippocampus 9: 495-499. |
| [36] |
Vargha-Khadem F, Gadian DG, Watkins KE, et al. (1997) Differential effects of early hippocampal pathology on episodic and semantic Memory. Science 277: 376-380. doi:10.1126/science.277.5324.376 doi: 10.1126/science.277.5324.376
|
| [37] | Douaud G, Groves AR, Tamnes CK, et al. (2014) A common brain network links development, aging, and vulnerability to disease. PNAS 111: 17649-17653. |
| [38] |
Miller EK, Fusi S (2013) Limber neurons for a nimble mind. Neuron 78: 211-213. doi:10.1016/j.neuron.2013.04.007. doi: 10.1016/j.neuron.2013.04.007
|
| [39] |
Rigotti M, Barak O, Warden MR, et al. (2013) The importance of mixed selectivity in complex cognitive tasks. Nature 497: 585-590. doi:10.1038/nature12160. doi: 10.1038/nature12160
|
| [40] |
Nestor PG, Nakamura M, Niznikiewicz M, et al. (2013) In search of the functional neuroanatomy of sociality: MRI subdivisions of orbital frontal cortex and social cognition. Soc Cogn Affective Neurosci 8: 460-467. doi:10.1039/scan/nss018 doi: 10.1093/scan/nss018
|
| [41] | Michael B, Spitzer, Robert L, et al. (2002) Structured Clinical Interview for DSM-IV-TR Axis I Disorders, Research Version, Non-patient Edition. (SCID-I/NP) New York: Biometrics Research, New York State Psychiatric Institute, November. |
| [42] | First MB, Gibbon M, Spitzer RL, et al. (1997) Structured Clinical Interview for DSM-IV Axis II Personality Disorders, (SCID-II). Washington, D.C.: American Psychiatric Press, Inc. |
| [43] |
Bouix S, Martin-Femandez M, Ungar L, et al. (2007) On evaluating brain tissue classifiers without a ground truth. NeuroImage 36: 1207-1224. doi: 10.1016/j.neuroimage.2007.04.031
|
| [44] |
Pohl K, Fisher J, Bouix S, et al. (2007) Using the logarithm of odds to define a vector space on probabilistic atlases. Med Image Analysis 11: 465-477. doi: 10.1016/j.media.2007.06.003
|
| [45] | Cohen J, Cohen P (1975) Applied multiple regression/correlation analysis of the behavioral sciences (2nd ed.). Hillsdale, NJ: Lawrence Erlbaum Associates. |
| [46] |
Cabeza R, Locantore JK, Anderson ND (2003) Lateralization of prefrontal activity during episodic memory retrieval: evidence for the production monitoring hypothesis. J Cogn Neurosci 15: 249-259. doi: 10.1162/089892903321208187
|
| [47] |
Reed JM, Squire LR (1997) Impaired recognition memory in patients with lesions limited to the hippocampal formation. Behavior Neurosci 111: 667-675. doi: 10.1037/0735-7044.111.4.667
|
| [48] |
Hwang K, Ghuman AS, Manoach DS, et al. (2014) Cortical neurodynamics of inhibitory control. J Neurosci 34: 9551-9561. doi: 10.1523/JNEUROSCI.4889-13.2014
|
| [49] |
Simon JS, Spiers HJ (2003) Prefrontal and medial temporal lobe interactions in long term memory. Nat Rev Neurosci 4: 637-648. doi: 10.1038/nrn1178
|
| [50] |
Nestor PG, Kubicki M, Kuroki N, et al. (2007) Episodic memory and neuroimaging of hippocampus and fornix in chronic schizophrenia. Psychiatry Res Neuroimaging 155: 21-28 doi: 10.1016/j.pscychresns.2006.12.020
|
| [51] |
Miller EK (2000) The prefrontal cortex and cognitive control. Nat Rev Neurosci 1: 59-65. doi: 10.1038/35036228
|
| [52] |
Shaw P, Greenstein D, Lerch J, et al. (2006) Intellectual ability and cortical development in children and adolescents. Nature 440: 676-679. doi: 10.1038/nature04513
|
| [53] |
Jung RE, Haier RJ (2007) The parieto-frontal integration theory (P-FIT): converging neuroimaging evidence. Behavior Brain Sci 30: 135-154. doi: 10.1017/S0140525X07001185
|
| [54] |
Kane MJ, Bleckley MK, Conway ARA, et al. (2001) A controlled attention view of working memory capacity. J Experiment Psychol General 130: 169-183. doi: 10.1037/0096-3445.130.2.169
|
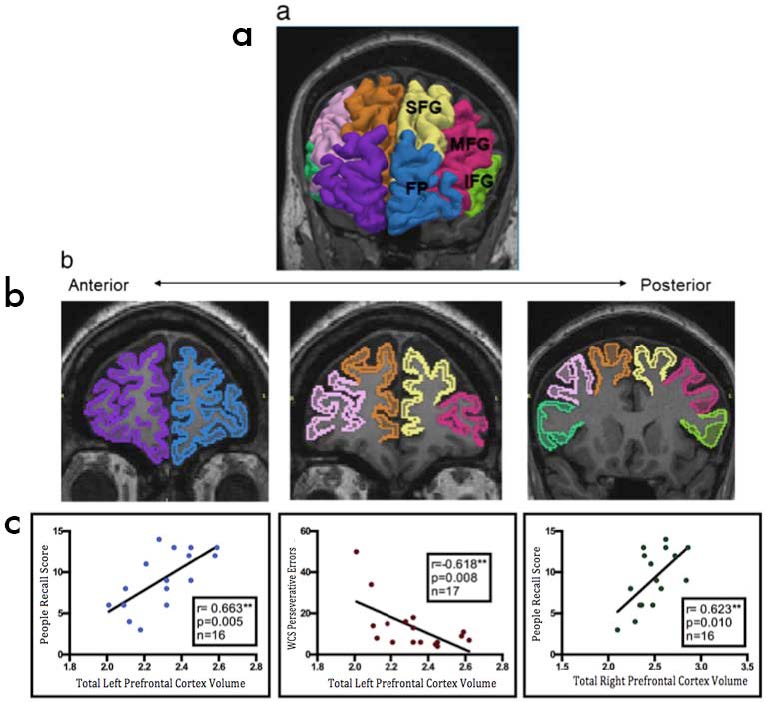
Figures(1) / Tables(4)

Paul G. Nestor, Toshiyuki Ohtani, James J. Levitt, Dominick T. Newell, Martha E. Shenton, Margaret Niznikiewicz, Robert W. McCarley. Prefrontal Lobe Gray Matter, Cognitive Control and Episodic Memory in Healthy Cognition[J]. AIMS Neuroscience, 2016, 3(3): 338-355. doi: 10.3934/Neuroscience.2016.3.338







 DownLoad:
DownLoad: