Citation: Hannah Nordberg, Michael J Hautus, Ernest Greene. Visual encoding of partial unknown shape boundaries[J]. AIMS Neuroscience, 2018, 5(2): 132-147. doi: 10.3934/Neuroscience.2018.2.132

| [1] | Fuchs W (1938) Untersuchung uber dae Sehen der Hemianopiker und Hemiamblyopiker [English title: Completion phenomena in hemianopic vision], In: Ellis W.D. Translator, A Source Book of Gestalt Psychology, London: Routledge & Kegan Paul, 352. |
| [2] |
Rosin P, Pantović J, Žunić J (2016) Measuring linearity of curves in 2D and 3D. Pattern Recognit 49: 65–78. doi: 10.1016/j.patcog.2015.07.011

|
| [3] |
Jomma HD, Hussein AI (2016) Circle views signature: A novel shape representation for shape recognition and retrieval. Can J Electr Comput Eng 39: 274–282. doi: 10.1109/CJECE.2016.2574745
|
| [4] |
Žunić J, Žunić D (2016) Shape interpretation of second-order moment invariants. J Math Imaging Vis 56: 125–136. doi: 10.1007/s10851-016-0638-8
|
| [5] |
Sharma S, Dubey S, Singh S, et al. (2015) Identity verification using shape and geometry of human hands. Expert Syst Appl Int J 42: 821–832. doi: 10.1016/j.eswa.2014.08.052
|
| [6] |
Tang K, Song P, Chen X (2017) 3D object recognition in cluttered scenes with robust shape description and correspondence selection. IEEE Access 5: 1833–1845. doi: 10.1109/ACCESS.2017.2658681
|
| [7] |
Sidram M, Bhajantri N (2015) An exploration with novel shape signature of GMSC distance function to track the object. Int J Imag Graph 15: 1550014. doi: 10.1142/S021946781550014X
|
| [8] |
Proenca H, Neves J, Barra S, et al. (2016) Joint head pose/soft label estimation for human recognition in-the-wild. IEEE Trans Pattern Anal Mach Intell 38: 2444–2456. doi: 10.1109/TPAMI.2016.2522441
|
| [9] | Tsai CY, Liao HC, Feng YC (2016) A novel translation, rotation, and scale-invariant shape description method for real-time speed-limit sign recognition. Int Conf Adv Mater Sci Eng 2017: 486–488. |
| [10] |
Greene E (2007) Retinal encoding of ultrabrief shape recognition cues. PloS One 2: e871. doi: 10.1371/journal.pone.0000871
|
| [11] | Greene E (2016) How do we know whether three dots form an equilateral triangle? JSM Brain Sci 1: 1002. |
| [12] | Greene E (2016) Retinal encoding of shape boundaries. JSM Anat Physiol 1: 1002. |
| [13] |
Greene E, Hautus MJ (2017) Demonstrating invariant encoding of shapes using a similarity judgment protocol. AIMS Neurosci 4: 120–146. doi: 10.3934/Neuroscience.2017.3.120
|
| [14] | Green DM, Swets JA (1966) Signal detection theory and psychophysics. New York: Wiley Press, 25: 1478–1481. |
| [15] |
Hautus MJ (1995) Corrections for extreme proportions and their biasing effects on estimated values of d′. Behav Res Methods Instrum Comput 27: 46–51. doi: 10.3758/BF03203619
|
| [16] |
Miller J (1996) The sampling distribution of d'. Percept Psychophys 58: 65–72. doi: 10.3758/BF03205476
|
| [17] |
Hautus M (1997) Calculating estimates of sensitivity from group data: Pooled versus averaged estimators. Behav Res Methods Instrum Comput 29: 556–562. doi: 10.3758/BF03210608
|
| [18] | Macmillan NA, Creelman CD (2005) Detection Theory: A User's Guide, 2 Eds., New Jersey: Lawrence Erlbaum. |
| [19] |
Hautus MJ, Hout DV, Lee HS (2009) Variants of a not-A and 2AFC tests: Signal detection theory models. Food Qual Prefer 20: 222–229. doi: 10.1016/j.foodqual.2008.10.002
|
| [20] | Hautus J (2012) SDT Assistant (version 1.0) [Software]. Available from: http://hautus.org. |
| [21] |
Greene E (2007) Recognition of objects displayed with incomplete sets of discrete boundary dots. Percept Mot Skills 104: 1043–1059. doi: 10.2466/pms.104.4.1043-1059
|
| [22] |
Sceniak MP, Hawken JJ, Shapley R (2001) Visual spatial characterization of macaque V1 neurons. J Neurophysiol 85: 1873–1887. doi: 10.1152/jn.2001.85.5.1873
|
| [23] |
Greene E (2008) Additional evidence that contour attributes are not essential cues for object recognition. Behav Brain Funct 4: e26. doi: 10.1186/1744-9081-4-26
|
| [24] |
Greene E, Ogden R (2012) Evaluating the contribution of shape attributes to recognition using the minimal transient discrete cue protocol. Behav Brain Funct 8: e53. doi: 10.1186/1744-9081-8-53
|
| [25] |
Fukushima K (1980) Neocognitron: A self-organizing neural network model for a mechanism of pattern recognition unaffected by shift in position. Biol Cybern 36: 193–202. doi: 10.1007/BF00344251
|
| [26] |
Rolls E, Cowey A, Bruce V (1992) Neurophysiological mechanisms underlying face processing within and beyond the temporal cortical visual areas. Phil Trans R Soc B 335: 11–21. doi: 10.1098/rstb.1992.0002
|
| [27] |
Wallis G, Rolls E (1997) Invariant face and object recognition in the visual system. Prog Neurobiol 51: 167–194. doi: 10.1016/S0301-0082(96)00054-8
|
| [28] |
Riesenhuber M, Poggio T (2000) Models of object recognition. Nat Neurosci 3: 1199–1204. doi: 10.1038/81479
|
| [29] | Suzuki N, Hashimoto N, Kashimori Y, et al. (2004) A neural model of predictive recognition in form pathway of visual cortex. Bio Syst 76: 33–42. |
| [30] |
Pinto N, Cox D, Dicarlo J (2008) Why is real-world visual object recognition hard? PLoS Comput Biol 4: e27. doi: 10.1371/journal.pcbi.0040027
|
| [31] |
Rodríguez-Sánchez A, Tsotsos J (2012) The roles of endstopped and curvature tuned computations in a hierarchical representation of 2D shape. PLoS One 7: e42058. doi: 10.1371/journal.pone.0042058
|
| [32] |
Hopfield J (1995) Pattern recognition computation using action potential timing for stimulus representation. Nature 376: 33–36. doi: 10.1038/376033a0
|
| [33] |
Thorpe S, Fize D, Marlot C (1996) Speed of processing in the human visual system. Nature 381: 520–522. doi: 10.1038/381520a0
|
| [34] |
Thorpe S, Delorme A, VanRullen R (2001) Spike-based strategies for rapid processing. Neural Netw 14: 715–725. doi: 10.1016/S0893-6080(01)00083-1
|
| [35] | VanRullen R, Thorpe S (2001) Is it a bird? Is it a plane? Ultra-rapid visual categorisation of natural and artifactual objects. Perception 30: 655–668. |
| [36] |
VanRullen R, Thorpe S (2002) Surfing a spike wave down the ventral stream. Vision Res 42: 2593–2615. doi: 10.1016/S0042-6989(02)00298-5
|
| [37] | Greene E, Patel Y (2018) Scan transcription of two-dimensional shapes as an alternative neuromorphic concept. Trends Artif Intell 1: 27–33. |
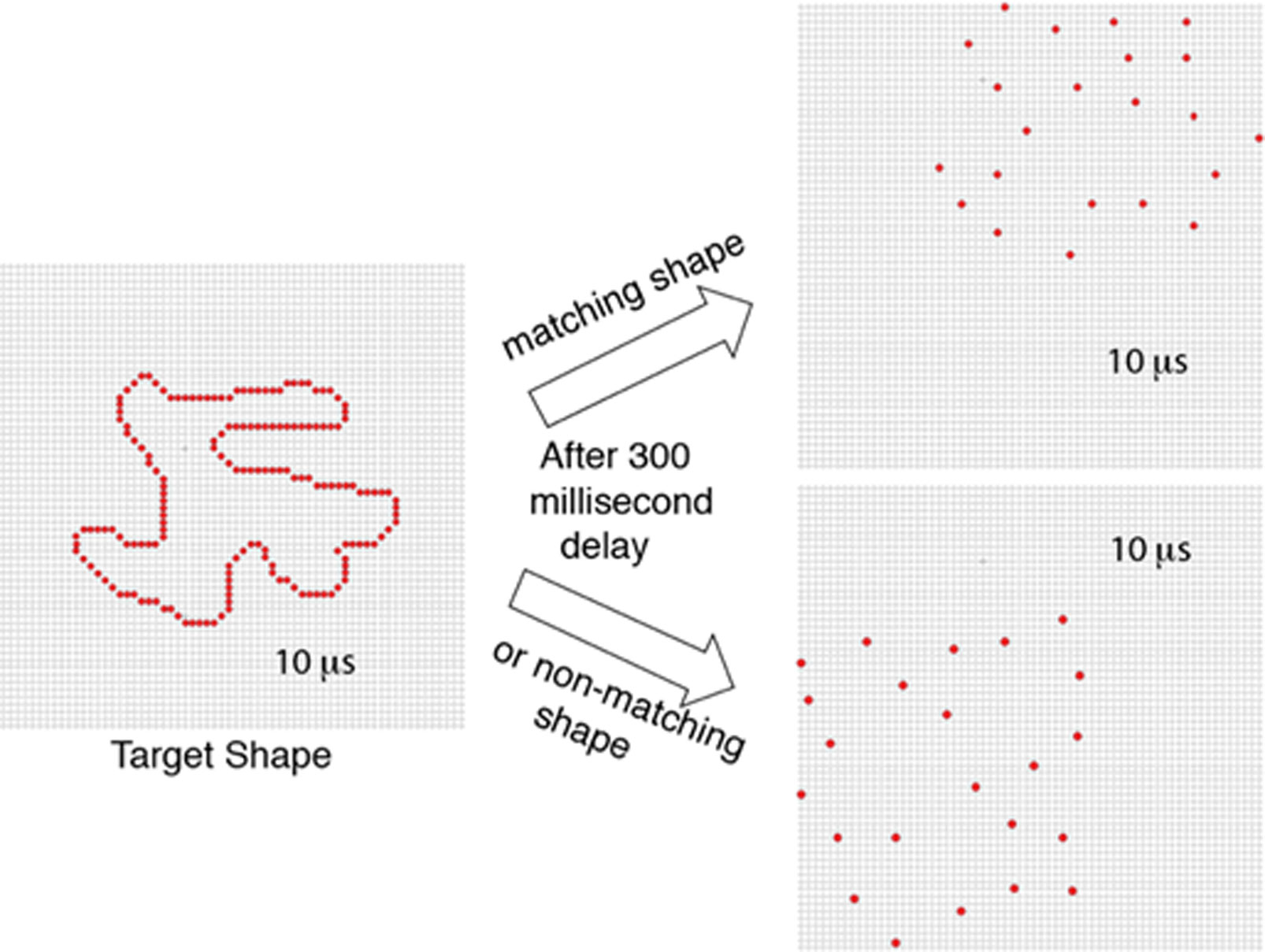
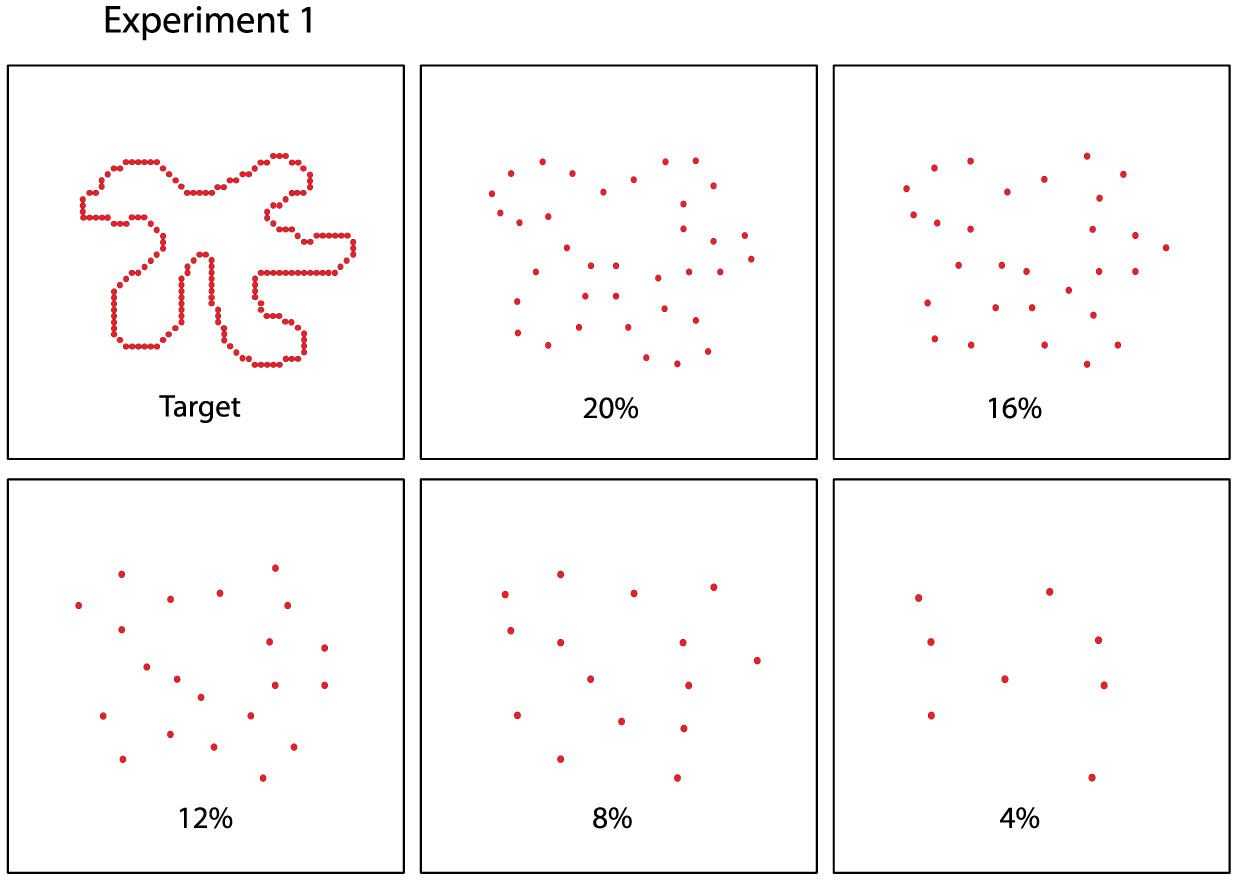
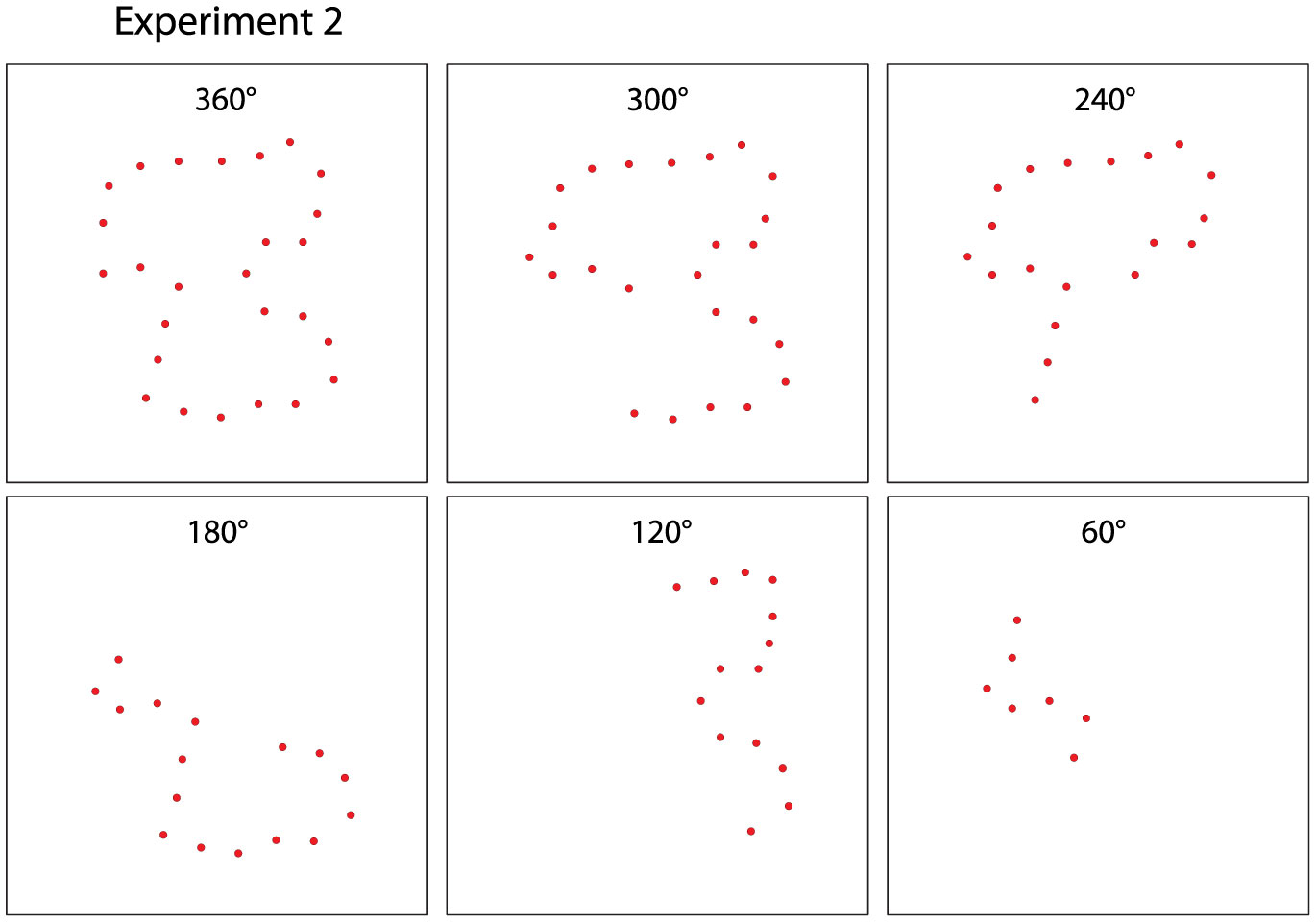
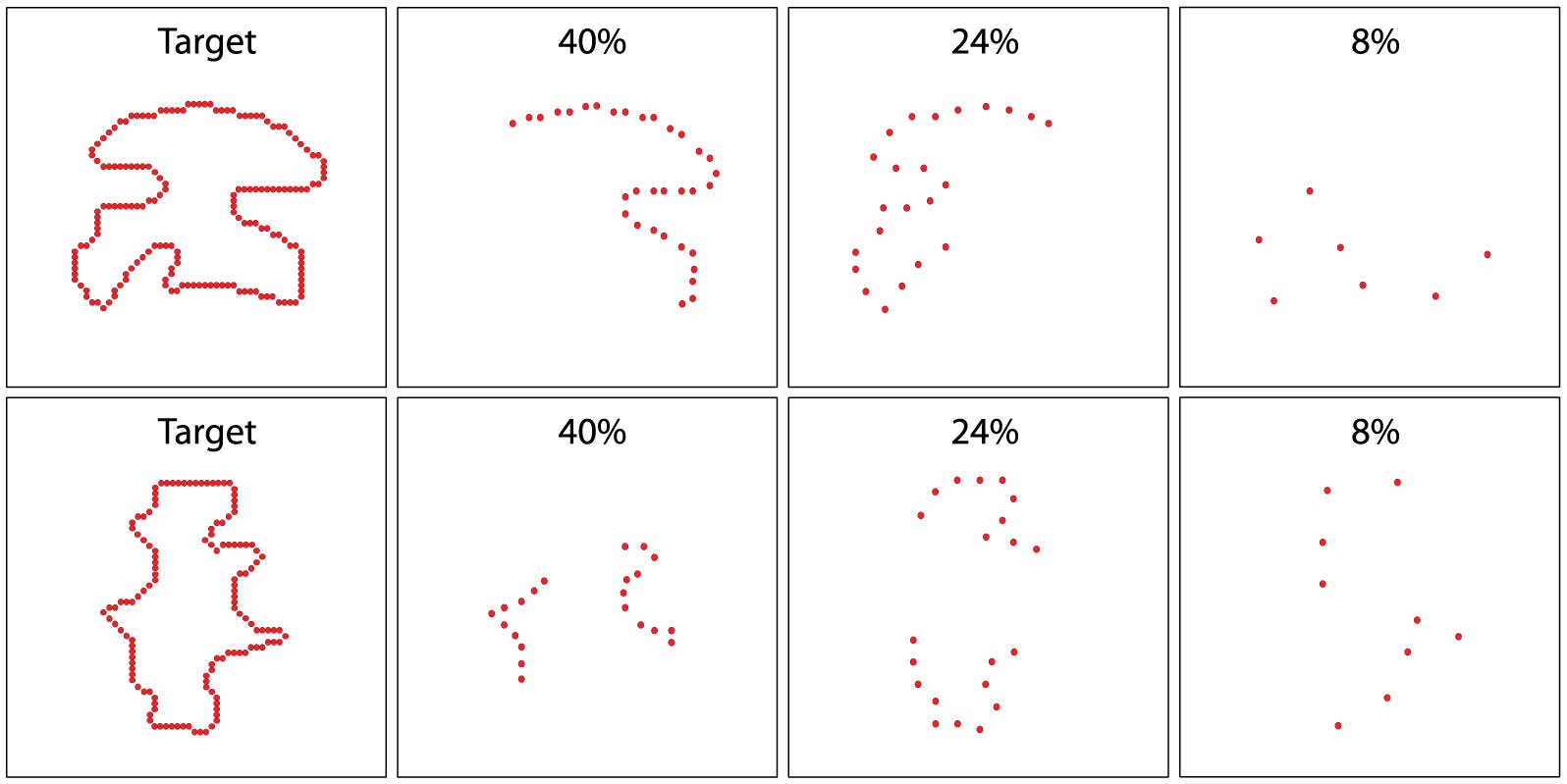
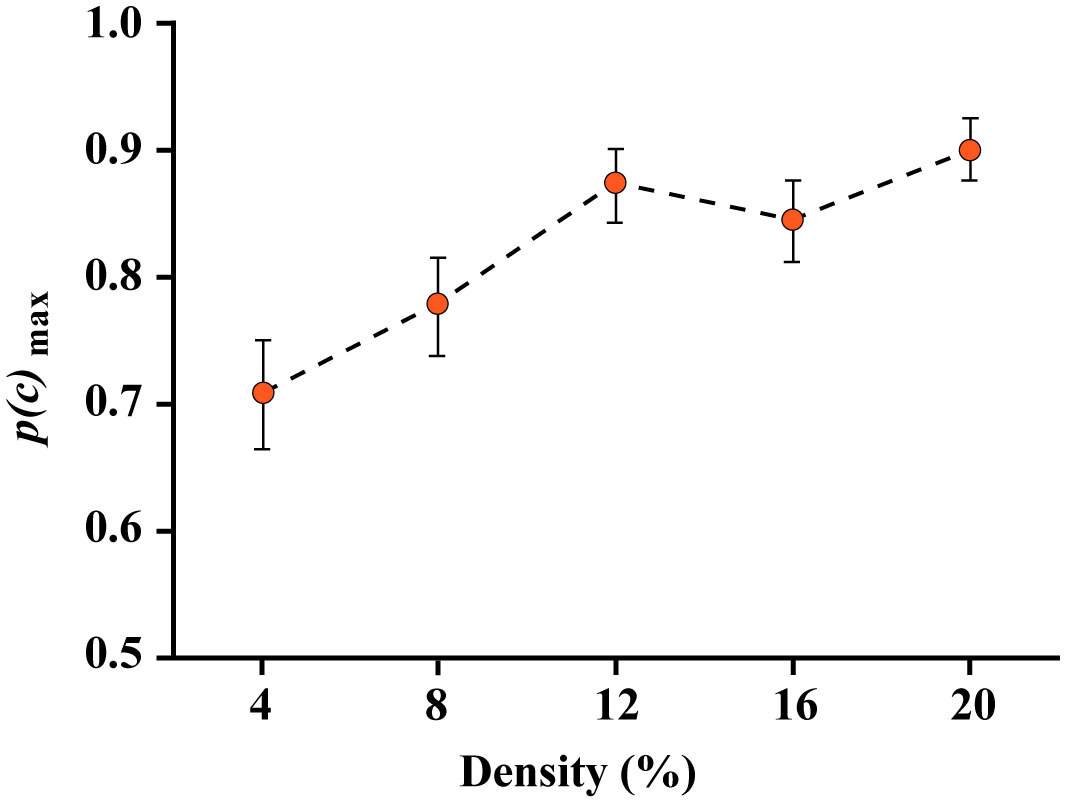
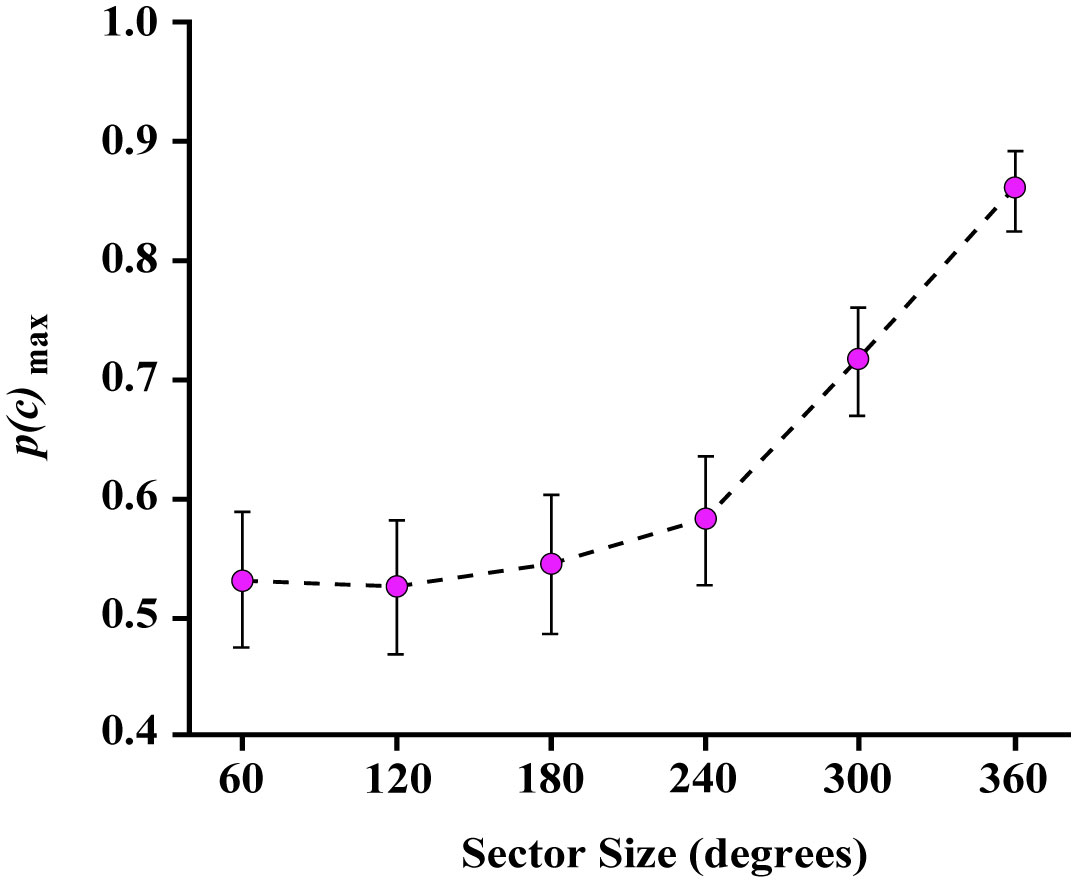
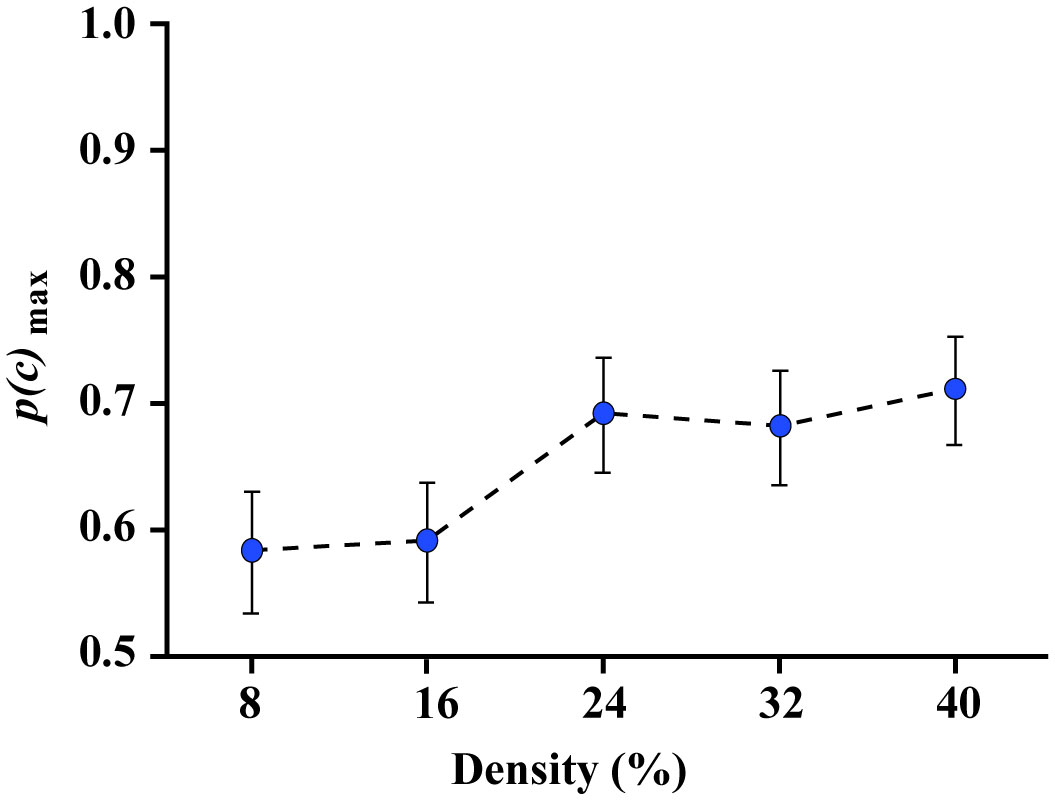
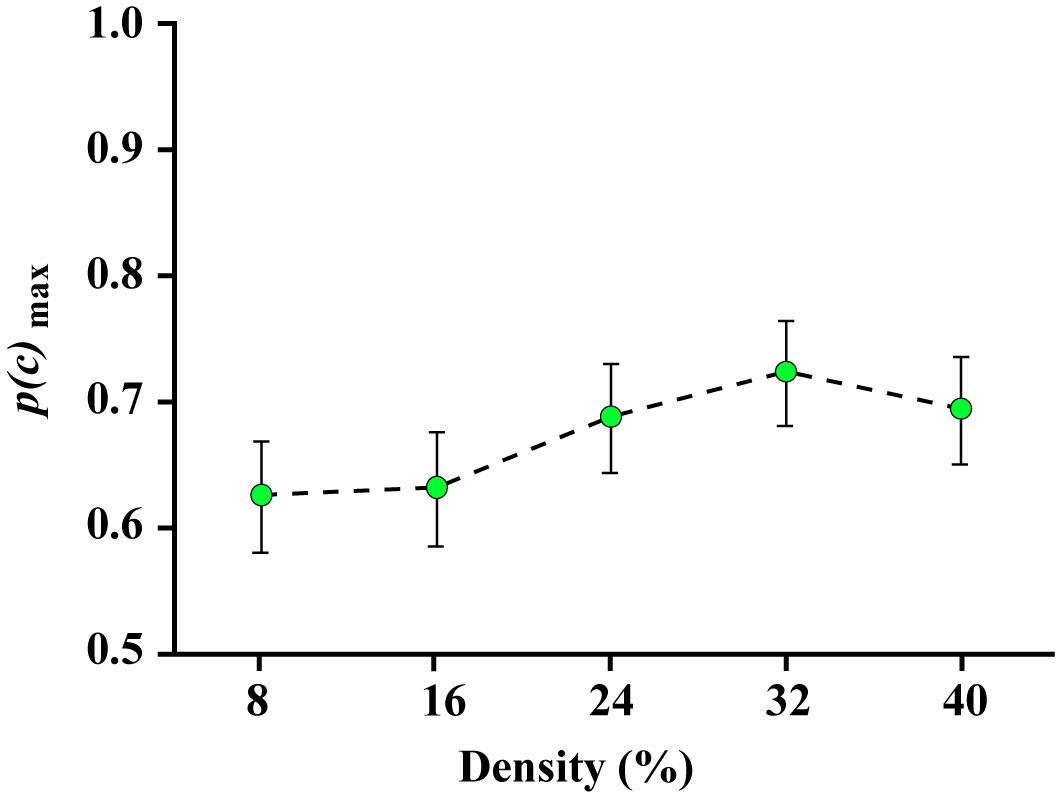
Figures(8)

Hannah Nordberg, Michael J Hautus, Ernest Greene. Visual encoding of partial unknown shape boundaries[J]. AIMS Neuroscience, 2018, 5(2): 132-147. doi: 10.3934/Neuroscience.2018.2.132







 DownLoad:
DownLoad: