Citation: Shizuka Takaku, Naoko Niimi, Toshihiko Kadoya, Hideji Yako, Masami Tsukamoto, Kunihiko Sakumi, Yusaku Nakabeppu, Hidenori Horie, Kazunori Sango. Galectin-1 and galectin-3 as key molecules for peripheral nerve degeneration and regeneration[J]. AIMS Molecular Science, 2016, 3(3): 325-337. doi: 10.3934/molsci.2016.3.325

| [1] |
Yang RY, Rabinovich GA, Liu FT (2008) Galectins: structure, function and therapeutic potential. Expert Rev Mol Med 10: e17. doi: 10.1017/S1462399408000719

|
| [2] | Sango K, Yanagisawa H, Watabe K, et al. (2012) Chapter 3: Galectin-1 as a multifunctional molecule in the peripheral nervous system after injury, In: Rayegani, S.M. Editor, Basic Principles of Peripheral Nerve Disorders, Rijeka, InTech d.o.o., 31-46. |
| [3] |
Takaku S, Yanagisawa H, Watabe K, et al. (2013) GDNF promotes neurite outgrowth and upregulates galectin-1 through the RET/PI3K signaling in cultured adult rat dorsal root ganglion neurons. Neurochem Int 62: 330-339. doi: 10.1016/j.neuint.2013.01.008
|
| [4] |
Tsukamoto M, Sango K, Niimi N, et al. (2015) Upregulation of galectin-3 in immortalized Schwann cells IFRS1 under diabetic conditions. Neurosci Res 92: 80-85. doi: 10.1016/j.neures.2014.11.008
|
| [5] |
Delacour D, Koch A, Jacob R (2009) The role of galectins in protein trafficking. Traffic 10: 1405-1413 doi: 10.1111/j.1600-0854.2009.00960.x
|
| [6] | Tracey BM, Feizi T, Abbott WM, et al. (1992) Subunit molecular mass assignment of 14,654 Da to the soluble beta-galactoside-binding lectin from bovine heart muscle and demonstration of intramolecular disulfide bonding associated with oxidative inactivation. J Biol Chem 267: 10342-10347. |
| [7] | Horie H, Inagaki Y, Sohma Y, et al. (1999) Galectin-1 regulates initial axonal growth in peripheral nerves after axotomy. J Neurosci 19: 9964-9974. |
| [8] |
Inagaki Y, Sohma Y, Horie H, et al. (2000) Oxidized galectin-1 promotes axonal regeneration in peripheral nerves but does not possess lectin properties. Eur J Biochem 267: 2955-2964. doi: 10.1046/j.1432-1033.2000.01311.x
|
| [9] |
Kadoya T, Horie H (2005) Structural and functional studies of galectin-1: a novel axonal regeneration-promoting activity for oxidized galectin-1. Curr Drug Targets 6: 375-383. doi: 10.2174/1389450054022007
|
| [10] | Camby I, Le Mercier M, Lefranc F, et al. (2006) Galectin-1: a small protein with major functions. Glycobiology 16: 137R-157R. |
| [11] |
Plachta N, Annaheim C, Bissière S, et al. (2007) Identification of a lectin causing the degeneration of neuronal processes using engineered embryonic stem cells. Nature Neurosci 10: 712-719. doi: 10.1038/nn1897
|
| [12] | Pesheva P, Kuklinski S, Schmitz B, et al. (1998) Galectin-3 promotes neural cell adhesion and neurite growth. J Neurosci Res 154: 639-654. |
| [13] | Chen H-L, Liao F, Lin T-N, et al. (2014) Chapter 24: Galectins and neuroinflammation, In: Yu, R.K. & Schengrund C.-L. Editors, Glycobiology of the Nervous System, Advances in Neurobiology 9, New York, Springer Science+Business Media, 517-542. |
| [14] |
Regan LJ, Dodd J, Barondes SH, et al. (1986) Selective expression of endogenous lactose-binding lectins and lactoseries glycoconjugates in subsets of rat sensory neurons. Proc Natl Acad Sci U S A 83: 2248-2252. doi: 10.1073/pnas.83.7.2248
|
| [15] | Fukaya K, Hasegawa M, Mashitani T, et al. (2003) Oxidized galectin-1 stimulates the migration of Schwann cells from both proximal and distal stumps of transected nerves and promotes axonal regeneration after peripheral nerve injury. J Neuropathol Exp Neurol 62: 162-172. |
| [16] |
Akazawa C, Nakamura Y, Sango K, et al. (2004) Distribution of the galectin-1 mRNA in the rat nervous system: its transient upregulation in rat facial motor neurons after facial nerve axotomy. Neuroscience 125: 171-178. doi: 10.1016/j.neuroscience.2004.01.034
|
| [17] | Park MJ, Chung K (1999) Endogenous lectin (RL-29) expression of the autonomic preganglionic neurons in the rat spinal cord. Anat Rec 254: 53-60. |
| [18] |
Salt TE, Hill RG (1983) Neurotransmitter candidates of somatosensory primary afferent fibers. Neuroscience 10: 1083-1103. doi: 10.1016/0306-4522(83)90101-X
|
| [19] |
Sango K, Tokashiki A, Ajiki K, et al. (2004) Synthesis, localization and externalization of galectin-1 in mature dorsal root ganglion neurons and Schwann cells. Eur J Neurosci 19: 55-64. doi: 10.1046/j.1460-9568.2003.03102.x
|
| [20] |
McGraw J, Gaudet AD, Oschipok LW, et al. (2005) Altered primary afferent anatomy and reduced thermal sensitivity in mice lacking galectin-1. Pain 114: 7-18. doi: 10.1016/j.pain.2004.10.009
|
| [21] |
Takasaki I, Taniguchi K, Komatsu F, et al. (2012) Contribution of spinal galectin-3 to acute herpetic allodynia in mice. Pain 153: 585-592. doi: 10.1016/j.pain.2011.11.022
|
| [22] |
Molliver DC, Wright DE, Leitner ML, et al. (1997) IB4-binding DRG neurons switch from NGF to GDNF dependence in early postnatal life. Neuron 19: 849-861. doi: 10.1016/S0896-6273(00)80966-6
|
| [23] |
Ernsberger U (2009) Role of neurotrophin signalling in the differentiation of neurons from dorsal root ganglia and sympathetic ganglia. Cell Tissue Res 336: 349-384. doi: 10.1007/s00441-009-0784-z
|
| [24] |
Imbe H, Okamoto K, Kadoya T, et al. (2003) Galectin-1 is involved in the potentiation of neuropathic pain in the dorsal horn. Brain Res 993: 72-83. doi: 10.1016/j.brainres.2003.08.064
|
| [25] |
McGraw J, Gaudet AD, Oschipok LW, et al. (2005) Regulation of neuronal and glial galectin-1 expression by peripheral and central axotomy of rat primary afferent neurons. Exp Neurol 195: 103-114. doi: 10.1016/j.expneurol.2005.04.004
|
| [26] |
McGraw J, Oschipok LW, Liu J, et al. (2004) Galectin-1 expression correlates with the regenerative potential of rubrospinal and spinal motoneurons. Neuroscience 128: 713-719. doi: 10.1016/j.neuroscience.2004.06.075
|
| [27] | Reichert F, Saada A, Rotshenker S (1994) Peripheral nerve injury induces Schwann cells to express two macrophage phenotypes: phagocytosis and the galactose-specific lectin MAC-2. J Neurosci 14: 3231-3245. |
| [28] | Mietto BS, Mostacada K, Martinez AM (2015) Neurotrauma and inflammation: CNS and PNS responses. Mediat Inflamm 2015: 251204. |
| [29] |
Dussor G, Koerber HR, Oaklander AL, et al. (2009) Nucleotide signaling and cutaneous mechanisms of pain transduction. Brain Res Rev 60: 24-35. doi: 10.1016/j.brainresrev.2008.12.013
|
| [30] |
Carmillo P, Dagø L, Day ES, et al. (2005) Glial cell line-derived neurotrophic factor (GDNF) receptor a-1 (GFR a1) is highly selective for GDNF versus artemin. Biochemistry 44: 2545-2554. doi: 10.1021/bi049247p
|
| [31] | Sango K, Yanagisawa H, Takaku S, et al. (2011) Immortalized adult rodent Schwann cells as in vitro models to study diabetic neuropathy. Exp Diabetes Res 2011:374943. |
| [32] |
Scheib J, Höke A (2013) Advances in peripheral nerve regeneration. Nat Rev Neurol 9: 668-676. doi: 10.1038/nrneurol.2013.227
|
| [33] |
Outenreath RL, Jones AL (1992) Influence of an endogenous lectin substrate on cultured dorsal root ganglion cells. J Neurocytol 21: 788-795. doi: 10.1007/BF01237904
|
| [34] | Mahanthappa NK, Cooper DN, Barondes SH, et al. (1994) Rat olfactory neurons can utilize the endogenous lectin, L-14, in a novel adhesion mechanism. Development 120: 1373-1384. |
| [35] |
Puche AC, Poirier F, Hair M, et al. (1996) Role of galectin-1 in the developing mouse olfactory system. Dev Biol 179: 274-287. doi: 10.1006/dbio.1996.0257
|
| [36] |
Miura T, Takahashi M, Horie H, et al. (2004) Galectin-1b, a natural monomeric form of galectin-1 lacking its six amino-terminal residues promotes axonal regeneration but not cell death. Cell Death Differ 11: 1076-1083. doi: 10.1038/sj.cdd.4401462
|
| [37] | Horie H, Kadoya T, Hikawa N, et al. (2004) Oxidized galectin-1 stimulates macrophages to promote axonal regeneration in peripheral nerves after axotomy. J Neurosci 24: 1873-1880. |
| [38] | Echigo Y, Sugiki H, Koizumi Y, et al. (2010) Activation of RAW264.7 macrophages by oxidized galectin-1. Immunol Lett 131: 19-23. |
| [39] | Kogawa Y, Nakajima K, Sasaguri K, et al. (2011) Oxidized galectin-1 reduces lipopolysaccharide-induced increase of proinflammatory cytokine mRNA in cultured macrophages. Clin Cosmet Investig Dent 19: 1-8. |
| [40] |
Gaudet AD, Leung M, Poirier F, et al. (2009) A role for galectin-1 in the immune response to peripheral nerve injury. Exp Neurol 220: 320-327. doi: 10.1016/j.expneurol.2009.09.007
|
| [41] |
Gaudet AD, Sweet DR, Polinski NK, et al. (2015) Galectin-1 in injured rat spinal cord: implications for macrophage phagocytosis and neural repair. Mol Cell Neurosci 64: 84-94. doi: 10.1016/j.mcn.2014.12.006
|
| [42] |
Narciso MS, Mietto Bde S, Marques SA, et al. (2009) Sciatic nerve regeneration is accelerated in galectin-3 knockout mice. Exp Neurol 217: 7-15. doi: 10.1016/j.expneurol.2009.01.008
|
| [43] |
Mietto BS, Jurgensen S, Alves L, et al. (2013) Lack of galectin-3 speeds Wallerian degeneration by altering TLR and pro-inflammatory cytokine expressions in injured sciatic nerve. Eur J Neurosci 37: 1682-1690. doi: 10.1111/ejn.12161
|
| [44] |
Rotshenker S (2009) The role of Galectin-3/MAC-2 in the activation of the innate-immune function of phagocytosis in microglia in injury and disease. J Mol Neurosci 39: 99-103. doi: 10.1007/s12031-009-9186-7
|
| [45] |
Gustavsson P, Linsmeier CE, Leffler H, et al. (2007) Galectin-3 inhibits Schwann cell proliferation in cultured sciatic nerve. Neuroreport 18: 669-673. doi: 10.1097/WNR.0b013e3280bef97b
|
| [46] |
Tucker BA, Rahimtula M, Mearow KM (2006) Laminin and growth factor receptor activation stimulates differential growth responses in subpopulations of adult DRG neurons. Eur J Neurosci 24: 676-690. doi: 10.1111/j.1460-9568.2006.04963.x
|
| [47] | Wang W, Park JW, Wang JL, et al. (2006) Immunoprecipitation of spliceosomal RNAs by antisera to galectin-1 and galectin-3. Nucleic Acids Res 34: 5166-5174. |
| [48] |
Yagihashi S, Mizukami H, Sugimoto K (2011) Mechanism of diabetic neuropathy: Where are we now and where to go? J Diabetes Investig 2: 18-32. doi: 10.1111/j.2040-1124.2010.00070.x
|
| [49] |
Mizisin AP (2014) Mechanisms of diabetic neuropathy: Schwann cells. Handb Clin Neurol 126: 401-428. doi: 10.1016/B978-0-444-53480-4.00029-1
|
| [50] | Pugliese G, Iacobini C, Pesce CM, et al. (2015) Galectin-3: an emerging all-out player in metabolic disorders and their complications. Glycobiology 25: 136-150. |
| [51] | Pugliese G, Pricci F, Iacobini C, et al. (2001) Accelerated diabetic glomerulopathy in galectin-3/AGE receptor 3 knockout mice. FASEB J 15: 2471-2479. |
| [52] |
Kanda A, Noda K, Saito W, et al. (2015) Aflibercept Traps Galectin-1, an Angiogenic Factor Associated with Diabetic Retinopathy. Sci Rep 5: 17946. doi: 10.1038/srep17946
|
| [53] |
Liu Y, Long L, Yuan F, et al. (2015) High glucose-induced Galectin-1 in human podocytes implicates the involvement of Galectin-1 in diabetic nephropathy. Cell Biol Int 39: 217-223. doi: 10.1002/cbin.10363
|
| [54] |
Mussche S, De Paepe B, Smet J, et al. (2012) Proteomic analysis in giant axonal neuropathy: new insights into disease mechanisms. Muscle Nerve 46: 246-256. doi: 10.1002/mus.23306
|
| [55] |
Franco PG, Pasquini LA, Pérez MJ, et al. (2015) Paving the way for adequate myelination: The contribution of galectin-3, transferrin and iron. FEBS Lett 589: 3388-3395. doi: 10.1016/j.febslet.2015.08.001
|
| [56] | Peng W (2012) Intravenous immunoglobulin treatment on anti-GM1 antibodies associated neuropathies inhibits cholera toxin and galectin-1 binding to ganglioside GM1. Immunol Lett 143: 146-151. |
| [57] | Bojarová P, Křen V (2016) Sugared biomaterial binding lectins: achievements and perspectives. Biomater Sci in press. |
| [58] |
Kadoya T, Oyanagi K, Kawakami E, et al. (2005) Oxidized galectin-1 advances the functional recovery after peripheral nerve injury. Neurosci Lett 380: 284-288. doi: 10.1016/j.neulet.2005.01.054
|
| [59] | Scott SA, Bugarcic A, Blanchard H (2009) Characterisation of oxidized recombinant human galectin-1. Protein Pept Lett 16: 1249-1255. |
| [60] | Guardia CM, Caramelo JJ, Trujillo M, et al. (2014) Structural basis of redox-dependent modulation of galectin-1 dynamics and function. Glycobiology 24: 428-441. |
| [61] |
Lin YT, Chen JS, Wu MH, et al. (2015) Galectin-1 accelerates wound healing by regulating the neuropilin-1/Smad3/NOX4 pathway and ROS production in myofibroblasts. J Invest Dermatol 135: 258-268. doi: 10.1038/jid.2014.288
|
| [62] | Pugliese G, Iacobini C, Pesce CM, et al. (2015) Galectin-3: an emerging all-out player in metabolic disorders and their complications. Glycobiology 25: 136-150. |
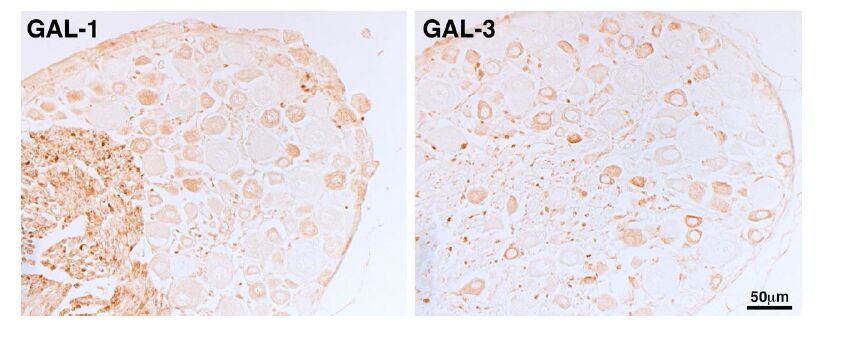
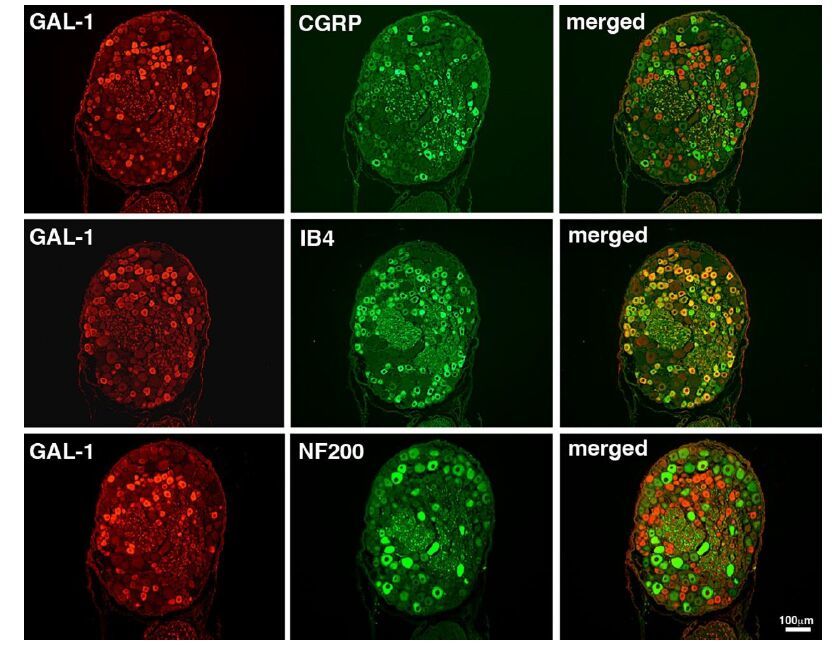
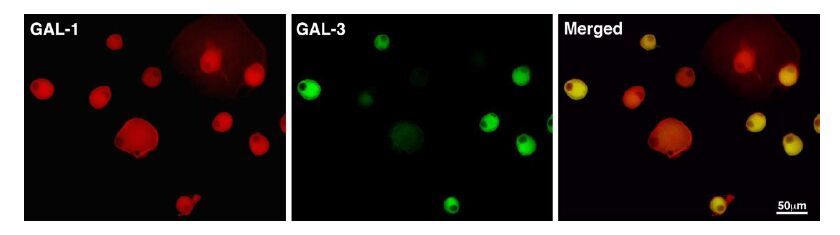
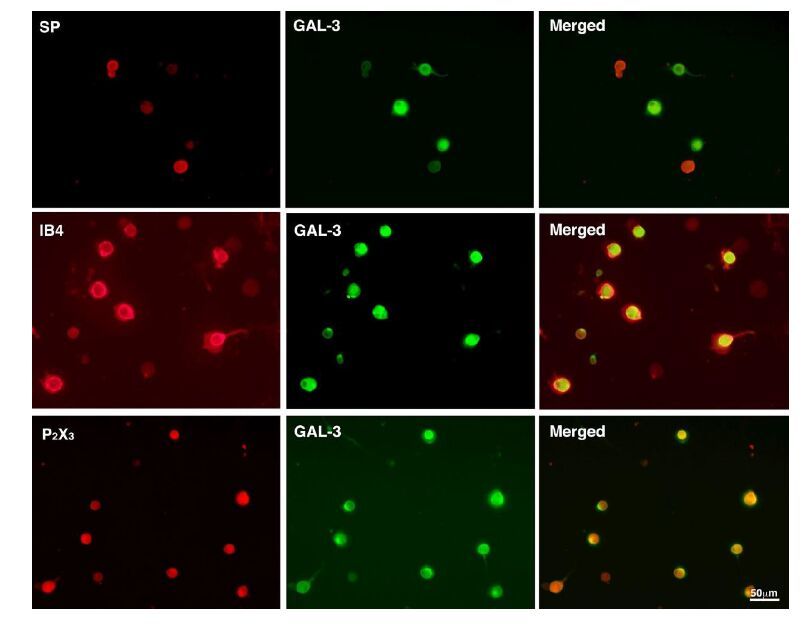
Figures(4)

Shizuka Takaku, Naoko Niimi, Toshihiko Kadoya, Hideji Yako, Masami Tsukamoto, Kunihiko Sakumi, Yusaku Nakabeppu, Hidenori Horie, Kazunori Sango. Galectin-1 and galectin-3 as key molecules for peripheral nerve degeneration and regeneration[J]. AIMS Molecular Science, 2016, 3(3): 325-337. doi: 10.3934/molsci.2016.3.325







 DownLoad:
DownLoad: