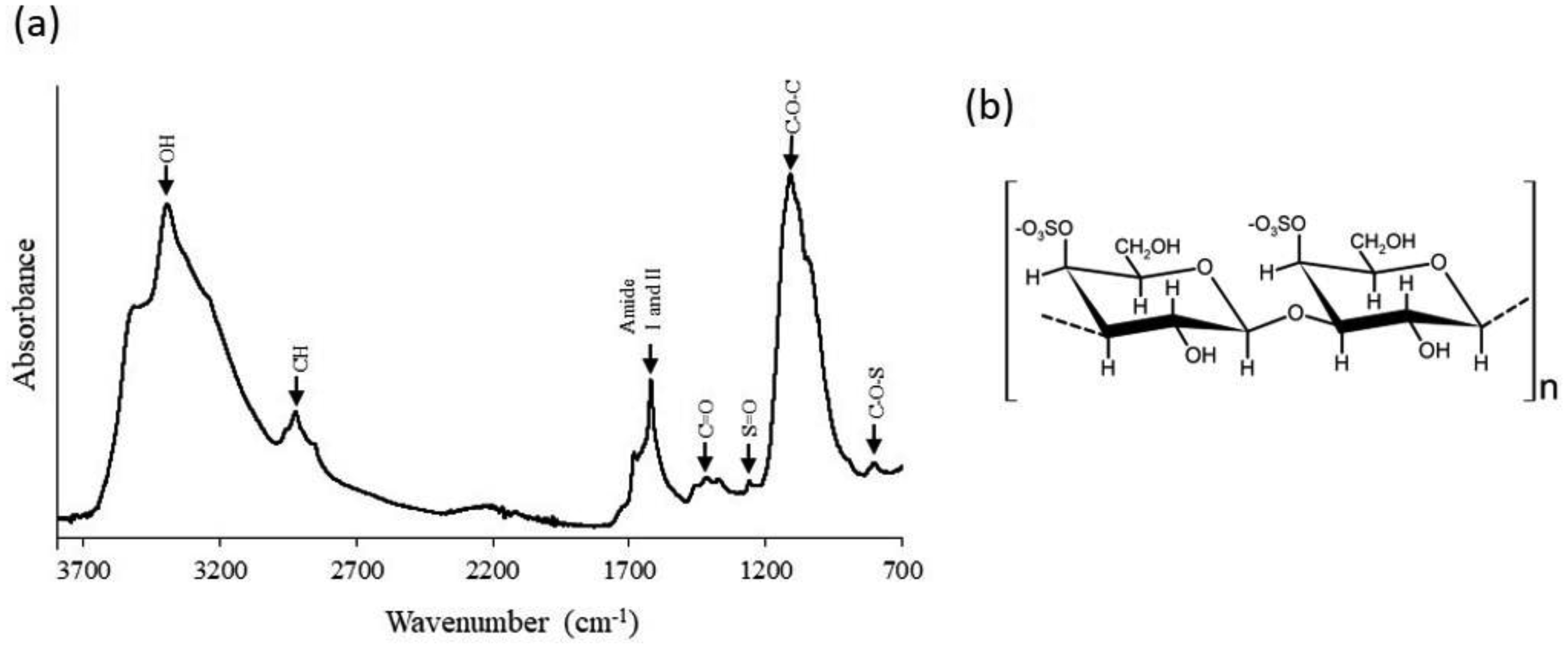
Increasing interest in biopolymers moves their knowledge frontiers. One area undergoing this development is polysaccharides. The practical and theoretical significance of studying polysaccharides lies in the numerous essential functions these macromolecules fulfill in living organisms, and the important industrial and technological purposes they serve. Polysaccharides are thought to be abundant in marine microalgae; nevertheless, little is known about their sulfated polysaccharides. We studied Chaetoceros muelleri sulfated polysaccharide (CMSP) chain conformation, physical characteristics, and morphology. The CMSP spectrum generated from Fourier-transform infrared analysis displayed distinctive bands for these macromolecules. S=O and C–O–S signals were detected at 1225 and 820 cm−1, respectively, confirming the presence of sulfate in the molecules. The biopolymer registered weight-average molar mass, intrinsic viscosity ([η]), radius of gyration (RG), hydrodynamic radius (Rh), and sulfate degree of substitution of 1933 kDa, 577 mL/g, 62 nm, 44 nm, and 0.5 (sulfates per disaccharide repeat unit), respectively. The exponent α and the coefficient K in the Mark-Houwink-Sakurada (MHS) equation were 0.76 and 9.76 x 10−3 mL/g, respectively. These values suggest a flexible and compact random coil structure in CMSP. The sample's zeta potential (ζ), conductivity, and diffusion coefficient (D) were −26.43 mV, −2.07 µm cm/s V, 1.25 mS/cm, and 1.8 × 10−8 cm2/s, correspondingly. The negative charge in the molecules is related to the sulfate groups contained. The CMSP surface was coarse and craggy, according to scanning electron microscopy (SEM). The information generated in this present study contributes to elucidating characteristics as fundamental knowledge to understand the macromolecule functionality.
Citation: Valeria Miranda-Arizmendi, Jorge Marquez-Escalante, Agustin Rascon-Chu, Karla Martínez-Robinson, Francisco Brown-Bojorquez, Elizabeth Carvajal-Millan. Chaetoceros muelleri sulfated polysaccharides: chain conformation, physical characteristics, and morphology[J]. AIMS Biophysics, 2024, 11(4): 455-463. doi: 10.3934/biophy.2024025
Increasing interest in biopolymers moves their knowledge frontiers. One area undergoing this development is polysaccharides. The practical and theoretical significance of studying polysaccharides lies in the numerous essential functions these macromolecules fulfill in living organisms, and the important industrial and technological purposes they serve. Polysaccharides are thought to be abundant in marine microalgae; nevertheless, little is known about their sulfated polysaccharides. We studied Chaetoceros muelleri sulfated polysaccharide (CMSP) chain conformation, physical characteristics, and morphology. The CMSP spectrum generated from Fourier-transform infrared analysis displayed distinctive bands for these macromolecules. S=O and C–O–S signals were detected at 1225 and 820 cm−1, respectively, confirming the presence of sulfate in the molecules. The biopolymer registered weight-average molar mass, intrinsic viscosity ([η]), radius of gyration (RG), hydrodynamic radius (Rh), and sulfate degree of substitution of 1933 kDa, 577 mL/g, 62 nm, 44 nm, and 0.5 (sulfates per disaccharide repeat unit), respectively. The exponent α and the coefficient K in the Mark-Houwink-Sakurada (MHS) equation were 0.76 and 9.76 x 10−3 mL/g, respectively. These values suggest a flexible and compact random coil structure in CMSP. The sample's zeta potential (ζ), conductivity, and diffusion coefficient (D) were −26.43 mV, −2.07 µm cm/s V, 1.25 mS/cm, and 1.8 × 10−8 cm2/s, correspondingly. The negative charge in the molecules is related to the sulfate groups contained. The CMSP surface was coarse and craggy, according to scanning electron microscopy (SEM). The information generated in this present study contributes to elucidating characteristics as fundamental knowledge to understand the macromolecule functionality.

| [1] |
Cai MH, Chen XY, Fu LQ, et al. (2021) Design and development of hybrid hydrogels for biomedical applications: recent trends in anticancer drug delivery and tissue engineering. Front Bioeng Biotech 9: 630943. https://doi.org/10.3389/fbioe.2021.630943 
|
| [2] |
Malafaya PB, Silva GA, Reis RL (2007) Natural–origin polymers as carriers and scaffolds for biomolecules and cell delivery in tissue engineering applications. Adv Drug Deliver Rev 59: 207-233. https://doi.org/10.1016/j.addr.2007.03.012
|
| [3] |
Kuznetsova TA, Andryukov BG, Besednova NN, et al. (2020) Marine algae polysaccharides as basis for wound dressings, drug delivery, and tissue engineering: a review. J Mar Sci Eng 8: 481. https://doi.org/10.3390/jmse8070481
|
| [4] |
Tiwari A, Melchor-Martínez EM, Saxena A, et al. (2021) Therapeutic attributes and applied aspects of biological macromolecules (polypeptides, fucoxanthin, sterols, fatty acids, polysaccharides, and polyphenols) rom diatoms—a review. Int J Biol Macromol 171: 398-413. https://doi.org/10.1016/j.ijbiomac.2020.12.219
|
| [5] |
Gilly W, Markaida U, Daniel P, et al. (2022) Long-term hydrographic changes in the gulf of California and ecological impacts: a crack in the world's aquarium?. Prog Oceanogr 206: 102857. https://doi.org/10.1016/j.pocean.2022.102857
|
| [6] | Miranda-Arizmendi V, Carvajal-Millan E, Fimbres-Olivarria D, et al. (2022) Macromolecular characteristics of sulfated extracellular polysaccharides from Chaetoceros muelleri. Proceedings of the 2nd International Electronic Conference on Diversity (IECD 2022)—New Insights into the Biodiversity of Plants, Animals and Microbes . Basel: Switzerland. https://doi.org/10.3390/IECD2022-12352 |
| [7] |
Li S, Zhao Z, He Z, et al. (2024) Effect of structural features on the antitumor activity of plant and microbial polysaccharides: a review. Food Biosci 61: 104648. https://doi.org/10.1016/j.fbio.2024.104648
|
| [8] |
Ptak SH, Hjuler AL, Ditlevsen SI, et al. (2021) The effect of seasonality and geographic location on sulphated polysaccharides from brown algae. Aquac Res 52: 6235-6243. https://doi.org/10.1111/are.15485
|
| [9] |
Fimbres-Olivarria D, Marquez-Escalante J, Martínez-Robinson KG, et al. (2023) Physicochemical and microstructural characteristics of sulfated polysaccharide from marine microalga. Analytica 4: 527-537. https://doi.org/10.3390/analytica4040036
|
| [10] |
Rochas C, Lahaye M, Yaphe W (1986) Sulfate content of carrageenan and agar determined by infrared spectroscopy. Bot Mar 29: 335-340. https://doi.org/10.1515/botm.1986.29.4.335
|
| [11] |
Miranda-Arizmendi V, Fimbres-Olivarria D, Miranda-Baeza A, et al. (2024) Sulfated polysaccharides from marine diatoms: insight into molecular characteristics and biological activity. AIMS Bioeng 11: 110-129. https://doi.org/10.3934/bioeng.2024007
|
| [12] |
Liu M, Wang Y, Wang R, et al. (2024) Preparation and performance evaluation of polysaccharide-iron complex of Eucommia ulmoides. Foods 22: 2302. https://doi.org/10.3390/foods13142302
|
| [13] |
Hong T, Yin JY, Nie SP, et al. (2021) Applications of infrared spectroscopy in polysaccharide structural analysis: progress, challenge and perspective. Food Chem: X 12: 100168. https://doi.org/10.1016/j.fochx.2021.100168
|
| [14] |
Wang Z, Wang L, Yu X, et al. (2024) Effect of polysaccharide addition on food physical properties: a review. Food Chem 431: 137099. https://doi.org/10.1016/j.foodchem.2023.137099
|
| [15] |
Li S, Zhao Z, He Z, et al. (2024) Effect of structural features on the antitumor activity of plant and microbial polysaccharides: a review. Food Biosci 61: 104648. https://doi.org/10.1016/j.fbio.2024.104648
|
| [16] |
Yang S, Wan H, Wang R, et al. (2019) Sulfated polysaccharides from Phaeodactylum tricornutum: isolation, structural characteristics, and inhibiting HEPG2 growth activity in vitro. PeerJ 7: e6409. https://doi.org/10.7717/peerj.6409
|
| [17] |
Goh KKT, Pinder DN, Hall CE, et al. (2006) Rheological and light scattering properties of flaxseed polysaccharide aqueous solutions. Biomacromolecules 7: 3098-3103. https://doi.org/10.1021/bm060577u
|
| [18] |
Picout DR, Ross-Murphy SB, Errington N, et al. (2003) Pressure cell assisted solubilization of xyloglucans: tamarind seed polysaccharide and detarium gum. Biomacromolecules 4: 799-807. https://doi.org/10.1021/bm0257659
|
| [19] |
Maciel B, Oelschlaeger C, Willenbacher N (2020) Chain flexibility and dynamics of alginate solutions in different solvents. Colloid Polym Sci 298: 791-801. https://doi.org/10.1007/s00396-020-04612-9
|
| [20] |
Hunter RJ (1981) The calculation of zeta potential. Zeta Potential in Colloid Science Principles and Applications . New York: Academic Press 59-124. https://doi.org/10.1016/C2013-0-07389-6
|
| [21] |
Wang Z, Luo D (2023) Extraction optimization, structure features, and bioactivities of two polysaccharides from Corydalis decumbens. Plos One 18: e0284413. https://doi.org/10.1371/journal.pone.0284413
|


Figures(3) / Tables(2)

Valeria Miranda-Arizmendi, Jorge Marquez-Escalante, Agustin Rascon-Chu, Karla Martínez-Robinson, Francisco Brown-Bojorquez, Elizabeth Carvajal-Millan. Chaetoceros muelleri sulfated polysaccharides: chain conformation, physical characteristics, and morphology[J]. AIMS Biophysics, 2024, 11(4): 455-463. doi: 10.3934/biophy.2024025







 DownLoad:
DownLoad: