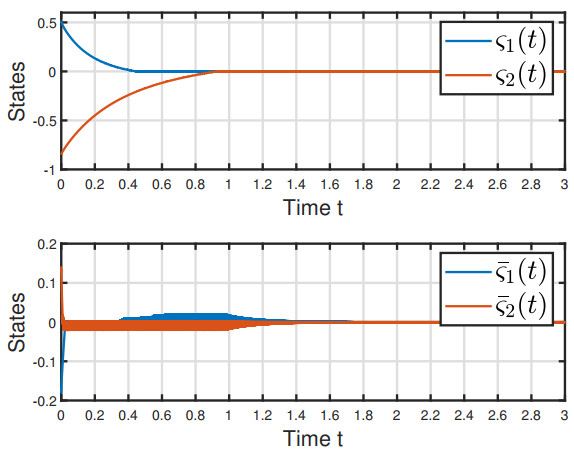
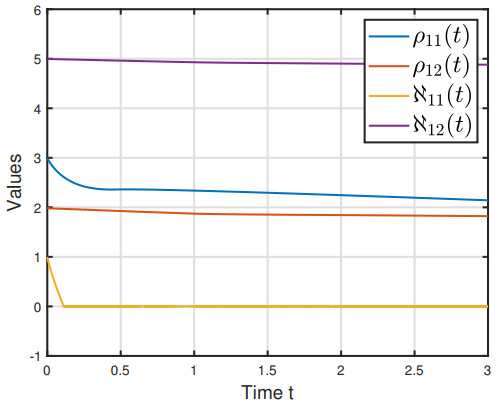
This paper explored the finite-time stability (FTS) of fractional-order fuzzy inertial neural network with mixed delays. First, the dimension of the model was reduced by the order reduction method. Second, by leveraging the fractional-order finite-time stability theorem, fractional calculus and inequality methods, we established some sufficient conditions to guarantee the FTS of the model under feasible delay-dependent feedback controller and delay-dependent adaptive controller, respectively. Additionally, we derived the settling times (STs) for each control strategy. Finally, we provided two examples to substantiate our findings.
Citation: Tiecheng Zhang, Liyan Wang, Yuan Zhang, Jiangtao Deng. Finite-time stability for fractional-order fuzzy neural network with mixed delays and inertial terms[J]. AIMS Mathematics, 2024, 9(7): 19176-19194. doi: 10.3934/math.2024935
This paper explored the finite-time stability (FTS) of fractional-order fuzzy inertial neural network with mixed delays. First, the dimension of the model was reduced by the order reduction method. Second, by leveraging the fractional-order finite-time stability theorem, fractional calculus and inequality methods, we established some sufficient conditions to guarantee the FTS of the model under feasible delay-dependent feedback controller and delay-dependent adaptive controller, respectively. Additionally, we derived the settling times (STs) for each control strategy. Finally, we provided two examples to substantiate our findings.

| [1] |
Y. Ni, Z. Wang, X. Huang, Q. Ma, H. Shen, Intermittent sampled-data control for local stabilization of neural networks subject to actuator saturation: A work-interval-dependent functional approach, IEEE Trans. Neural Netw. Learn. Syst., 35 (2024), 1087–1097. https://doi.org/10.1109/TNNLS.2022.3180076 doi: 10.1109/TNNLS.2022.3180076

|
| [2] |
H. B. Zeng, Y. He, K. L. Teo, Monotone-delay-interval-based Lyapunov functionals for stability analysis of systems with a periodically varying delay, Automatica, 138 (2022), 110030, 2022. https://doi.org/10.1016/j.automatica.2021.110030 doi: 10.1016/j.automatica.2021.110030
|
| [3] |
Y. Sheng, F. L. Lewis, Z. Zeng, T. Huang, Lagrange stability and finite-time stabilization of fuzzy memristive neural networks with hybrid time-varying delays, IEEE Trans. Cybernet., 50 (2020), 2959–2970. https://doi.org/10.1109/TCYB.2019.2912890 doi: 10.1109/TCYB.2019.2912890
|
| [4] |
S. Ding, Z. Wang, Synchronization of coupled neural networks via an event-dependent intermittent pinning control, IEEE Trans. Syst. Man Cybernet., 52 (2022) 1928–1934. https://doi.org/10.1109/TSMC.2020.3035173 doi: 10.1109/TSMC.2020.3035173
|
| [5] |
Y. Wu, Y. Gao, W. Li, Finite-time synchronization of switched neural networks with state-dependent switching via intermittent control, Neurocomputing, 384 (2020), 325–334. https://doi.org/10.1016/j.neucom.2019.12.031 doi: 10.1016/j.neucom.2019.12.031
|
| [6] |
M. J. A. Schuetz, J. K. Brubaker, H. G. Katzgraber, Combinatorial optimization with physics-inspired graph neural networks, Nat. Mach. Intell., 4 (2022), 367–377. https://doi.org/10.1038/s42256-022-00468-6 doi: 10.1038/s42256-022-00468-6
|
| [7] |
A. Sellami, S. Tabbone, Deep neural networks-based relevant latent representation learning for hyperspectral image classification, Pattern Recogn., 121 (2022), 108224. https://doi.org/10.1016/j.patcog.2021.108224 doi: 10.1016/j.patcog.2021.108224
|
| [8] |
S. Panghal, M. Kumar, Optimization free neural network approach for solving ordinary and partial differential equations, Eng. Comput., 37 (2021), 2989–3002. https://doi.org/10.1007/s00366-020-00985-1 doi: 10.1007/s00366-020-00985-1
|
| [9] |
S. Wen, Z. Zeng, T. Huang, Q. Meng, W. Yao, Lag synchronization of switched neural networks via neural activation function and applications in image encryption, IEEE Trans. Neural Netw. Learn. Syst., 26 (2015), 1493–1502. https://doi.org/10.1109/TNNLS.2014.2387355 doi: 10.1109/TNNLS.2014.2387355
|
| [10] |
K. L. Babcock, R. M. Westervelt, Stability and dynamics of simple electronic neural networks with added inertia, Phys. D, 23 (1986), 464–469. https://doi.org/10.1016/0167-2789(86)90152-1 doi: 10.1016/0167-2789(86)90152-1
|
| [11] |
K. L. Babcock, R. M. Westervelt, Dynamics of simple electronic neural networks, Phys. D, 28 (1987), 305–316. https://doi.org/10.1016/0167-2789(87)90021-2 doi: 10.1016/0167-2789(87)90021-2
|
| [12] |
Q. Cui, L. Li, J. Cao, Stability of inertial delayed neural networks with stochastic delayed impulses via matrix measure method, Neurocomputing, 471 (2022), 70–78. https://doi.org/10.1016/j.neucom.2021.10.113 doi: 10.1016/j.neucom.2021.10.113
|
| [13] |
C. Huang, Exponential stability of inertial neural networks involving proportional delays and non-reduced order method, J. Exp. Theor. Artif. In., 32 (2020), 133–146. https://doi.org/10.1080/0952813X.2019.1635654 doi: 10.1080/0952813X.2019.1635654
|
| [14] |
S. Dong, H. Zhu, S. Zhong, K. Shi, Y. Liu, New study on fixed-time synchronization control of delayed inertial memristive neural networks, Appl. Math. Comput., 399 (2021), 126035. https://doi.org/10.1016/j.amc.2021.126035 doi: 10.1016/j.amc.2021.126035
|
| [15] |
L. Ke, Mittag-Leffler stability and asymptotic $\omega$-periodicity of fractional-order inertial neural networks with time-delays, Neurocomputing, 465 (2021), 53–62. https://doi.org/10.1016/j.neucom.2021.08.121 doi: 10.1016/j.neucom.2021.08.121
|
| [16] |
F. Wei, G. Chen, Z. Zeng, N. Gunasekaran, Finite/fixed-time synchronization of inertial memristive neural networks by interval matrix method for secure communication, Neural Netw., 167 (2023), 168–182. https://doi.org/10.1016/j.neunet.2023.08.015 doi: 10.1016/j.neunet.2023.08.015
|
| [17] |
C. Li, W. Deng, Remarks on fractional derivatives, Appl. Math. Comput., 187 (2007), 777–784. https://doi.org/10.1016/j.amc.2006.08.163 doi: 10.1016/j.amc.2006.08.163
|
| [18] |
Y. Gu, H. Wang, Y. Yu, Stability and synchronization for Riemann-Liouville fractional-order time-delayed inertial neural networks, Neurocomputing, 340 (2019), 270–280. https://doi.org/10.1016/j.neucom.2019.03.005 doi: 10.1016/j.neucom.2019.03.005
|
| [19] |
L. Luo, L. Li, W. Huang, Q. Cui, Stability of the Caputo fractional-order inertial neural network with delay-dependent impulses, Neurocomputing, 520 (2023), 25–32. https://doi.org/10.1016/j.neucom.2022.11.060 doi: 10.1016/j.neucom.2022.11.060
|
| [20] |
Q. Peng, J. Jian, Synchronization analysis of fractional-order inertial-type neural networks with time delays, Math. Comput. Simul., 205 (2023), 62–77. https://doi.org/10.1016/j.matcom.2022.09.023 doi: 10.1016/j.matcom.2022.09.023
|
| [21] |
Y. Liu, Y. Sun, L. Liu, Stability analysis and synchronization control of fractional-order inertial neural networks with time-varying delay, IEEE Access, 10 (2022), 56081–56093. https://doi.org/10.1109/ACCESS.2022.3178123 doi: 10.1109/ACCESS.2022.3178123
|
| [22] |
G. Rajchakit, R. Sriraman, Robust passivity and stability analysis of uncertain complex-valued impulsive neural networks with time-varying delays, Neural Process Lett., 53 (2021), 581–606. https://doi.org/10.1007/s11063-020-10401-w doi: 10.1007/s11063-020-10401-w
|
| [23] |
F. Kong, Q. Zhu, R. Sakthivel, A. Mohammadzadeh, Fixed-time synchronization analysis for discontinuous fuzzy inertial neural networks with parameter uncertainties, Neurocomputing, 422 (2021), 295–313. https://doi.org/10.1016/j.neucom.2020.09.014 doi: 10.1016/j.neucom.2020.09.014
|
| [24] |
S. Wen, G. Bao, Z. Zeng, Y. Chen, T. Huang, Global exponential synchronization of memristor-based recurrent neural networks with time-varying delays, Neural Netw., 48 (2013), 195–203. https://doi.org/10.1016/j.neunet.2013.10.001 doi: 10.1016/j.neunet.2013.10.001
|
| [25] |
F. Du, J. G. Lu, Finite-time stability of fractional-order fuzzy cellular neural networks with time delays, Fuzzy Sets Syst., 438 (2022), 107–120. https://doi.org/10.1016/j.fss.2021.08.011 doi: 10.1016/j.fss.2021.08.011
|
| [26] |
F. Yan, D. Luo, Finite-time stability of Caputo fractional fuzzy differential equations with delay in granular sense, Commun. Nonlinear Sci. Numer. Simul., 134 (2024), 108022. https://doi.org/10.1016/j.cnsns.2024.108022 doi: 10.1016/j.cnsns.2024.108022
|
| [27] |
B. Zheng, Z. Wang, Mittag-Leffler synchronization of fractional-order coupled neural networks with mixed delays, Appl. Math. Comput., 430 (2022), 127303. https://doi.org/10.1016/j.amc.2022.127303 doi: 10.1016/j.amc.2022.127303
|
| [28] |
S. Wen, M. Z. Q. Chen, Z. Zeng, T. Huang, C. Li, Adaptive neural-fuzzy sliding-mode fault-tolerant control for uncertain nonlinear systems, IEEE Trans. Syst. Man Cybernet., 47 (2017), 2268–2278. https://doi.org/10.1109/TSMC.2017.2648826 doi: 10.1109/TSMC.2017.2648826
|
| [29] | T. Yang, L. Yang, C. W. Wu, L. O. Chua, Fuzzy cellular neural networks: theory, In: 1996 Fourth IEEE international workshop on cellular neural networks and their applications proceedings (CNNA-96), IEEE, 1996,181–186. https://doi.org/10.1109/CNNA.1996.566545 |
| [30] |
C. Aouiti, J. Cao, H. Jallouli, C. Huang, Finite-time stabilization for fractional-order inertial neural networks with time varying delays, Nonlinear Anal. Model. Control, 27 (2022), 1–18. https://doi.org/10.15388/namc.2022.27.25184 doi: 10.15388/namc.2022.27.25184
|
| [31] |
T. Hu, Z. He, X. Zhang, S. Zhong, Finite-time stability for fractional-order complex-valued neural networks with time delay, Appl. Math. Comput., 365 (2020), 124715. https://doi.org/10.1016/j.amc.2019.124715 doi: 10.1016/j.amc.2019.124715
|
| [32] |
H. L. Li, C. Hu, L. Zhang, H. Jiang, J. Cao, Complete and finite-time synchronization of fractional-order fuzzy neural networks via nonlinear feedback control, Fuzzy Sets Syst., 443 (2022), 50–69. https://doi.org/10.1016/j.fss.2021.11.004 doi: 10.1016/j.fss.2021.11.004
|
| [33] |
F. Du, J. Lu, Finite-time synchronization of fractional-order delayed fuzzy cellular neural networks with parameter uncertainties, IEEE Trans. Fuzzy Syst., 31 (2023), 1769–1779. https://doi.org/10.1109/TFUZZ.2022.3214070 doi: 10.1109/TFUZZ.2022.3214070
|
| [34] |
L. Ke, Mittag-Leffler stability and asymptotic $\omega$-periodicity of fractional-order inertial neural networks with time-delays, Neurocomputing, 465 (2021), 53–62. https://doi.org/10.1016/j.neucom.2021.08.121 doi: 10.1016/j.neucom.2021.08.121
|
| [35] |
E. Arslan, G. Narayanan, M. S. Ali, S. Arik, S. Saroha, Controller design for finite-time and fixed-time stabilization of fractional-order memristive complex-valued BAM neural networks with uncertain parameters and time-varying delays, Neural Netw., 130 (2020), 60–74. https://doi.org/10.1016/j.neunet.2020.06.021 doi: 10.1016/j.neunet.2020.06.021
|
| [36] |
M. Zheng, L. Li, H. Peng, J. Xiao, Y. Yang, Y. Zhang, et al., Finite-time stability and synchronization of memristor-based fractional-order fuzzy cellular neural networks, Commun. Nonlinear Sci. Numer. Simul., 59 (2018), 272–291. https://doi.org/10.1016/j.cnsns.2017.11.025 doi: 10.1016/j.cnsns.2017.11.025
|
| [37] |
A. Polyakov, Nonlinear feedback design for fixed-time stabilization of linear control systems, IEEE Trans. Automat. Control, 57 (2012), 2106–2110. https://doi.org/10.1109/TAC.2011.2179869 doi: 10.1109/TAC.2011.2179869
|
| [38] |
Y. Li, Y. Kao, C. Wang, H. Xia, Finite-time synchronization of delayed fractional-order heterogeneous complex networks, Neurocomputing, 384 (2020), 368–375. https://doi.org/10.1016/j.neucom.2019.11.043 doi: 10.1016/j.neucom.2019.11.043
|
| [39] |
C. Aouiti, Q. Hui, H. Jallouli, E. Moulay, Fixed-time stabilization of fuzzy neutral-type inertial neural networks with time-varying delay, Fuzzy Sets Syst., 411 (2021), 48–67. https://doi.org/10.1016/j.fss.2020.10.018 doi: 10.1016/j.fss.2020.10.018
|
| [40] |
Y. Liu, G. Zhang, J. Hu, Fixed-time stabilization and synchronization for fuzzy inertial neural networks with bounded distributed delays and discontinuous activation functions, Neurocomputing, 495 (2022), 86–96. https://doi.org/10.1016/j.neucom.2022.04.101 doi: 10.1016/j.neucom.2022.04.101
|
| [41] |
C. Aouiti, H. Jallouli, State feedback controllers based finite-time and fixed-time stabilisation of high order BAM with reaction-diffusion term, Int. J. Syst. Sci., 52 (2021), 905–927. https://doi.org/10.1080/00207721.2020.1849861 doi: 10.1080/00207721.2020.1849861
|
| [42] |
G. Zhang, Z. Zeng, Stabilization of second-order memristive neural networks with mixed time delays via nonreduced order, IEEE Trans. Neural Netw. Learn. Syst., 31 (2020), 700–706. https://doi.org/10.1109/TNNLS.2019.2910125 doi: 10.1109/TNNLS.2019.2910125
|
| [43] |
G. Zhang, J. Cao, New results on fixed/predefined-time synchronization of delayed fuzzy inertial discontinuous neural networks: Non-reduced order approach, Appl. Math. Comput., 440 (2023), 127671. https://doi.org/10.1016/j.amc.2022.127671 doi: 10.1016/j.amc.2022.127671
|
| [44] |
J. Yu, C. Hu, H. Jiang, L. Wang, Exponential and adaptive synchronization of inertial complex-valued neural networks: A non-reduced order and non-separation approach, Neural Netw., 124 (2020), 50–59. https://doi.org/10.1016/j.neunet.2020.01.002 doi: 10.1016/j.neunet.2020.01.002
|
| [45] |
J. Wang, X. Wang, Y. Wang, X. Zhang, Non-reduced order method to global h-stability criteria for proportional delay high-order inertial neural networks, Appl. Math. Comput., 407 (2021), 126308. https://doi.org/10.1016/j.amc.2021.126308 doi: 10.1016/j.amc.2021.126308
|
| [46] |
J. Jian, L. Duan, Finite-time synchronization for fuzzy neutral-type inertial neural networks with time-varying coefficients and proportional delays, Fuzzy Sets Syst., 381 (2020), 51–67. https://doi.org/10.1016/j.fss.2019.04.004 doi: 10.1016/j.fss.2019.04.004
|
| [47] |
M. Zheng, L. Li, H. Peng, J. Xiao, Y. Yang, Y. Zhang, et al., Finite-time stability and synchronization of memristor-based fractional-order fuzzy cellular neural networks, Commun. Nonlinear Sci. Numer. Simul., 59 (2018), 272–291. https://doi.org/10.1016/j.cnsns.2017.11.025 doi: 10.1016/j.cnsns.2017.11.025
|
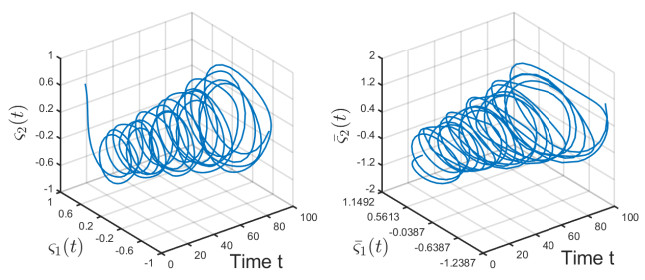
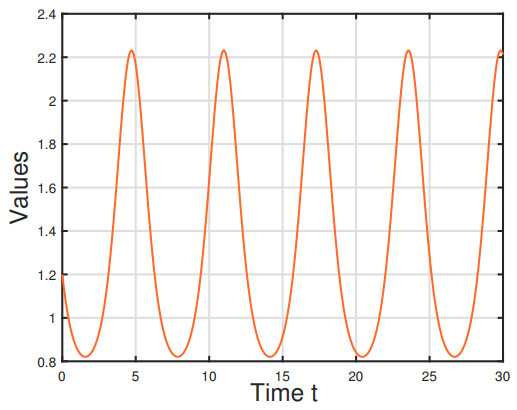
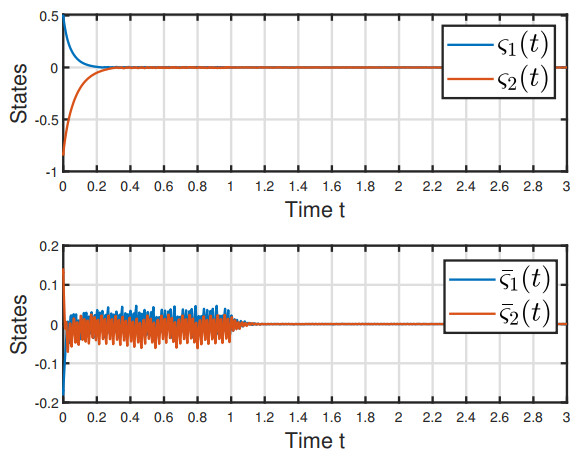
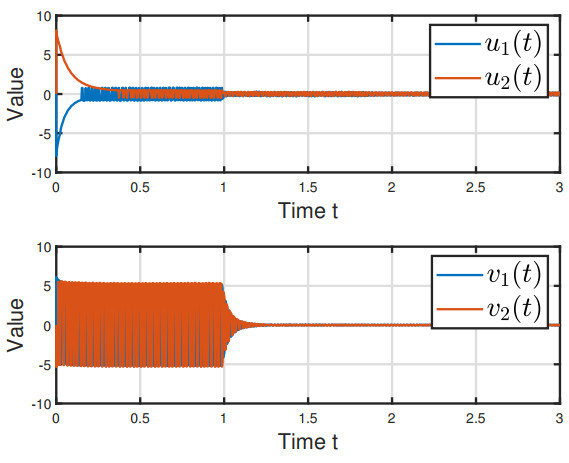
Figures(6)

Tiecheng Zhang, Liyan Wang, Yuan Zhang, Jiangtao Deng. Finite-time stability for fractional-order fuzzy neural network with mixed delays and inertial terms[J]. AIMS Mathematics, 2024, 9(7): 19176-19194. doi: 10.3934/math.2024935







 DownLoad:
DownLoad: