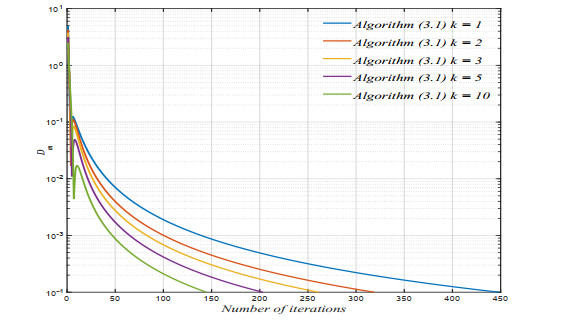
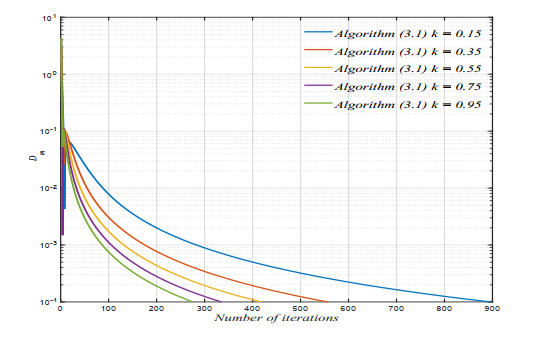
The purpose of this paper was to prove that a novel algorithm with an inertial approach, used to generate an iterative sequence, strongly converges to a fixed point of a nonexpansive mapping in a real uniformly convex Banach space with a uniformly Gâteaux differentiable norm. Furthermore, zeros of accretive mappings were obtained. The proposed algorithm has been implemented and tested via numerical simulation in MATLAB. The simulation results showed that the algorithm converges to the optimal configurations and shows the effectiveness of the proposed algorithm.
Citation: Kaiwich Baewnoi, Damrongsak Yambangwai, Tanakit Thianwan. A novel algorithm with an inertial technique for fixed points of nonexpansive mappings and zeros of accretive operators in Banach spaces[J]. AIMS Mathematics, 2024, 9(3): 6424-6444. doi: 10.3934/math.2024313
The purpose of this paper was to prove that a novel algorithm with an inertial approach, used to generate an iterative sequence, strongly converges to a fixed point of a nonexpansive mapping in a real uniformly convex Banach space with a uniformly Gâteaux differentiable norm. Furthermore, zeros of accretive mappings were obtained. The proposed algorithm has been implemented and tested via numerical simulation in MATLAB. The simulation results showed that the algorithm converges to the optimal configurations and shows the effectiveness of the proposed algorithm.

| [1] | C. E. Chidume, Geometric properties of Banach spaces and nonlinear iterations, Lecture Notes in Mathematics 1965, Springer, 2009. https://doi.org/10.1007/978-1-84882-190-3 |
| [2] | R. P. Agarwal, D. O'Regan, D. R. Sahu, Fixed point theory for Lipschtz-type mappings with applications, Berlin: Springer, 2008. |
| [3] | W. Takahashi, Nonlinear functional analysis: Fixed point theory and its applications, Yokohama: Yokohama Publishers, 2000. |
| [4] | V. Berinde, Iterative approximation of fixed points, Lectures Notes 1912, Springer, 2002. |
| [5] |
F. E. Browder, Nonexpansive nonlinear operators in Banach spaces, P. Nat. Acad. Sci., 54 (1965), 1041–1044. https://doi.org/10.1073/pnas.54.4.1041 doi: 10.1073/pnas.54.4.1041

|
| [6] |
Z. Opial, Weak convergence of successive approximations for nonexpansive mappings, B. Am. Math. Soc., 73 (1967), 591–597. https://doi.org/10.1090/S0002-9904-1967-11761-0 doi: 10.1090/S0002-9904-1967-11761-0
|
| [7] | M. A. Krasnosel'skii, Two remarks on the method of successive approximations, Uspekhi Mat. Nauk, 10 (1955), 123–127. |
| [8] |
W. R. Mann, Mean value methods in iteration, P. Am. Math. Soc., 4 (1953), 506–510. https://doi.org/10.1090/S0002-9939-1953-0054846-3 doi: 10.1090/S0002-9939-1953-0054846-3
|
| [9] |
C. W. Groetsch, A note on segmenting Mann iterates, J. Math. Anal. Appl., 40 (1972), 369–372. https://doi.org/10.1016/0022-247X(72)90056-X doi: 10.1016/0022-247X(72)90056-X
|
| [10] |
T. L. Hicks, J. D. Kubicek, On the Mann iteration process in a Hilbert space, J. Math. Anal. Appl., 59 (1977), 498–504. https://doi.org/10.1016/0022-247X(77)90076-2 doi: 10.1016/0022-247X(77)90076-2
|
| [11] |
B. P. Hillam, A generalization of Krasnoselski's theorem on the real line, Math. Mag., 48 (1975), 167–168. https://doi.org/10.1080/0025570X.1975.11976471 doi: 10.1080/0025570X.1975.11976471
|
| [12] |
M. Edelstein, R. C. O'Brien, Nonexpansive mappings, asymptotic regularity and successive approximations, J. Lond. Math. Soc., 2 (1978), 547–554. https://doi.org/10.1112/jlms/s2-17.3.547 doi: 10.1112/jlms/s2-17.3.547
|
| [13] |
M. Bravo, R. Cominetti, M. P. Signé, Rates of convergence for inexact Krasnosel'skii-Mann iterations in Banach spaces, Math. Program., 175 (2019), 241–262. https://doi.org/10.1007/s10107-018-1240-1 doi: 10.1007/s10107-018-1240-1
|
| [14] |
Q. L. Dong, J. Huang, X. H. Li, Y. J. Cho, Th. M. Rassias, MiKM: Multi-step inertial Krasnosel'skii-Mann algorithm and its applications, J. Global Optim., 73 (2019), 801–824. https://doi.org/10.1007/s10898-018-0727-x doi: 10.1007/s10898-018-0727-x
|
| [15] |
Q. L. Dong, X. H. Li, Y. J. Cho, T. M. Rassias, Multi-step inertial Krasnosel'skii-Mann iteration with new inertial parameters arrays, J. Fix. Point Theory A., 23 (2021), 1–18. https://doi.org/10.1007/s11784-021-00879-9 doi: 10.1007/s11784-021-00879-9
|
| [16] |
S. He, Q. L. Dong, H. Tian, X. H. Li, On the optimal parameters of Krasnosel'skii-Mann iteration, Optimization, 70 (2021), 1959–1986. https://doi.org/10.1080/02331934.2020.1767101 doi: 10.1080/02331934.2020.1767101
|
| [17] | Q. L. Dong, Y. J. Cho, S. He, P. M. Pardalos, T. M. Rassias, The Krasnosel'skii-Mann iterative method: Recent progress and applications, Springer, 2022. https://doi.org/10.1007/978-3-030-91654-1 |
| [18] |
F. E. Browder, W. V. Petryshyn, The solution by iteration of nonlinear functional equations in Banach spaces, B. Am. Math. Soc., 72 (1966), 571–575. https://doi.org/10.1090/S0002-9904-1966-11544-6 doi: 10.1090/S0002-9904-1966-11544-6
|
| [19] | H. H. Bauschke, P. L. Combettes, Convex analysis and monotone operator theory in Hilbert spaces, 2 Eds., CMS Books in Mathematics, New York: Springer, 2017. https://doi.org/10.1007/978-3-319-48311-5 |
| [20] |
J. Borwein, S. Reich, I. Shafrir, Krasnoselski-Mann iterations in normed spaces, Can. Math. Bull., 35 (1992), 21–28. https://doi.org/10.4153/CMB-1992-003-0 doi: 10.4153/CMB-1992-003-0
|
| [21] |
S. Ishikawa, Fixed points and iteration of a nonexpansive mapping in a Banach space, P. Am. Math. Soc., 59 (1976), 65–71. https://doi.org/10.1090/S0002-9939-1976-0412909-X doi: 10.1090/S0002-9939-1976-0412909-X
|
| [22] |
S. Reich, Weak convergence theorems for nonexpansive mappings in Banach spaces, J. Math. Anal. Appl., 67 (1979), 274–276. https://doi.org/10.1016/0022-247X(79)90024-6 doi: 10.1016/0022-247X(79)90024-6
|
| [23] |
A. Genel, J. Lindenstrass, An example concerning fixed points, Isr. J. Math., 22 (1975), 81–86. https://doi.org/10.1007/BF02757276 doi: 10.1007/BF02757276
|
| [24] |
R. I. Bot, E. R. Csetnek, D. Meier, Inducing strong convergence into the asymptotic behavior of proximal splitting algorithms in Hilbert spaces, Optim. Method. Softw., 34 (2019), 489–514. https://doi.org/10.1080/10556788.2018.1457151 doi: 10.1080/10556788.2018.1457151
|
| [25] |
Q. L. Dong, Y. Y. Lu, J. Yang, The extragradient algorithm with inertial effects for solving the variational inequality, Optimization, 65 (2016), 2217–2226. https://doi.org/10.1080/02331934.2016.1239266 doi: 10.1080/02331934.2016.1239266
|
| [26] |
J. Fan, L. Liu, X. Qin, A subgradient extragradient algorithm with inertial effects for solving strongly pseudomonotone variational inequalities, Optimization, 69 (2020), 2199–2215. https://doi.org/10.1080/02331934.2019.1625355 doi: 10.1080/02331934.2019.1625355
|
| [27] |
B. T. Polyak, Some methods of speeding up the convergence of iteration methods, USSR Comput. Math. Math. Phys., 4 (1964), 1–17. https://doi.org/10.1016/0041-5553(64)90137-5 doi: 10.1016/0041-5553(64)90137-5
|
| [28] |
Q. L. Dong, H. B. Yuan, Y. J. Cho, T. M. Rassias, Modified inertial Mann algorithm and inertial CQ-algorithm for nonexpansive mappings, Optim. Lett., 12 (2018), 87–102. https://doi.org/10.1007/s11590-016-1102-9 doi: 10.1007/s11590-016-1102-9
|
| [29] |
H. A. Hammad, H. ur Rehman, M. De la Sen, Advanced algorithms and common solutions to variational inequalities, Symmetry, 12 (2020), 1198. https://doi.org/10.3390/sym12071198 doi: 10.3390/sym12071198
|
| [30] |
P. E. Maingé, Convergence theorems for inertial KM-type algorithms, J. Comput. Appl. Math., 219 (2008), 223–236. https://doi.org/10.1016/j.cam.2007.07.021 doi: 10.1016/j.cam.2007.07.021
|
| [31] |
Y. Shehu, X. H. Li, Q. L. Dong, An efficient projection-type method for monotone variational inequalities in Hilbert spaces, Numer. Algorithms, 84 (2020), 365–388. https://doi.org/10.1007/s11075-019-00758-y doi: 10.1007/s11075-019-00758-y
|
| [32] | B. Tan, S. Xu, S. Li, Inertial shrinking projection algorithms for solving hierarchical variational inequality problems, J. Nonlinear Convex A., 21 (2020), 871–884. |
| [33] | L. Liu, B. Tan, S. Y. Cho, On the resolution of variational inequality problems with a double-hierarchical structure, J. Nonlinear Convex A., 21 (2020), 377–386. |
| [34] | F. Akutsah, O. K. Narain, J. K. Kim, Improved generalized M-iteration for quasi-nonexpansive multivalued mappings with application in real Hilbert spaces, Nonlinear Funct. Anal. Appl., 27 (2022), 59–82. |
| [35] | N. D. Truong, J. K. Kim, T. H. H. Anh, Hybrid inertial contraction projection methods extended to variational inequality problems, Nonlinear Funct. Anal. Appl., 27 (2022), 203–221. |
| [36] |
J. A. Abuchu, G. C. Ugunnadi, O. K. Narain, Inertial proximal and contraction methods for solving monotone variational inclusion and fixed point problems, Nonlinear Funct. Anal. Appl., 28 (2023), 175–203. https://doi.org/10.23952/jnfa.2023.19 doi: 10.23952/jnfa.2023.19
|
| [37] | I. Cioranescu, Geometry of Banach spaces, duality mappings and nonlinear problems, Dordrecht: Kluwer Academic, 1990. https://doi.org/10.1007/978-94-009-2121-4 |
| [38] |
F. E. Browder, Nonlinear mappings of nonexpansive and accretive type in Banach spaces, B. Am. Math. Soc., 73 (1967), 875–882. https://doi.org/10.1090/S0002-9904-1967-11823-8 doi: 10.1090/S0002-9904-1967-11823-8
|
| [39] |
T. Kato, Nonlinear semigroups and evolution equations, J. Math. Soc. Jpn., 19 (1967), 508–520. https://doi.org/10.2969/jmsj/01940508 doi: 10.2969/jmsj/01940508
|
| [40] |
W. O. Ray, An elementary proof of surjectivity for a class of accretive operators, P. Am. Math. Soc., 75 (1979), 255–258. https://doi.org/10.1090/S0002-9939-1979-0532146-0 doi: 10.1090/S0002-9939-1979-0532146-0
|
| [41] | J. V. Caristi, The fixed point theory for mappings satisfying inwardness conditions, Ph.D. Thesis, The University of Iowa, Iowa City, 1975. |
| [42] |
H. Robert, J. Martin, Nonlinear operators and differential equations in Banach spaces, SIAM Rev., 20 (1978), 202–204. https://doi.org/10.1137/1020032 doi: 10.1137/1020032
|
| [43] |
R. H. Martin, A global existence theorem for autonomous differential equations in Banach spaces, P. Am. Math. Soc., 26 (1970), 307–314. https://doi.org/10.1090/S0002-9939-1970-0264195-6 doi: 10.1090/S0002-9939-1970-0264195-6
|
| [44] | F. E. Browder, Nonlinear elliptic boundary value problems, B. Am. Math. Soc., 69 (1963). https://doi.org/10.1090/S0002-9904-1963-11068-X |
| [45] | K. Deimling, Nonlinear functional analysis, Berlin: Springer, 1985. https://doi.org/10.1007/978-3-662-00547-7 |
| [46] | L. Wei, Q. Zhang, Y. Zhang, R. P. Agarwal, Iterative algorithm for zero points of the sum of countable accretive-type mappings and variational inequalities, J. Nonlinear Funct. Anal., 2022 (2022). https://doi.org/10.23952/jnfa.2022.3 |
| [47] |
H. K. Xu, N. Altwaijry, I. Alzughaibi, S. Chebbi, The viscosity approximation method for accretive operators in Banach spaces, J. Nonlinear Var. Anal., 6 (2022), 37–50. https://doi.org/10.23952/jnva.6.2022.1.03 doi: 10.23952/jnva.6.2022.1.03
|
| [48] |
W. L. Bynum, Normal structure coefficients for Banach spaces, Pac. J. Math., 86 (1980), 427–436. https://doi.org/10.2140/pjm.1980.86.427 doi: 10.2140/pjm.1980.86.427
|
| [49] |
T. C. Lim, H. K. Xu, Fixed point theorems for asymptotically nonexpansive mappings, Nonlinear Anal. TMA, 22 (1994), 1345–1355. https://doi.org/10.1016/0362-546X(94)90116-3 doi: 10.1016/0362-546X(94)90116-3
|
| [50] |
H. K. Xu, Inequalities in Banach spaces with applications, Nonlinear Anal. TMA, 16 (1991), 1127–1138. https://doi.org/10.1016/0362-546X(91)90200-K doi: 10.1016/0362-546X(91)90200-K
|
| [51] |
S. Shioji, W. Takahashim, Strong convergence of approximated sequences for nonexpansive mappings in Banach spaces, P. Am. Math. Soc., 125 (1997), 3641–3645. https://doi.org/10.1090/S0002-9939-97-04033-1 doi: 10.1090/S0002-9939-97-04033-1
|
| [52] |
H. K. Xu, Iterative algorithms for nonlinear operators, J. London Math. Soc., 66 (2002), 240–256. https://doi.org/10.1112/S0024610702003332 doi: 10.1112/S0024610702003332
|
| [53] |
W. Shatanawi, A. Bataihah, A. Tallafha, Four-step iteration scheme to approximate fixed point for weak contractions, CMC-Comput. Mater. Con., 64 (2020), 1491–1504. https://doi.org/10.32604/cmc.2020.010365 doi: 10.32604/cmc.2020.010365
|
Figures(9) / Tables(2)

Kaiwich Baewnoi, Damrongsak Yambangwai, Tanakit Thianwan. A novel algorithm with an inertial technique for fixed points of nonexpansive mappings and zeros of accretive operators in Banach spaces[J]. AIMS Mathematics, 2024, 9(3): 6424-6444. doi: 10.3934/math.2024313







 DownLoad:
DownLoad: