Citation: Bindu Subhadra, Man Hwan Oh, Chul Hee Choi. Quorum sensing in Acinetobacter: with special emphasis on antibiotic resistance, biofilm formation and quorum quenching[J]. AIMS Microbiology, 2016, 2(1): 27-41. doi: 10.3934/microbiol.2016.1.27

| [1] |
Forster DH, Daschner FD (1998) Acinetobacter species as nosocomial pathogens. Eur J Clin Microbiol Infect Dis 17: 73–77. doi: 10.1007/BF01682159

|
| [2] |
Cisneros JM, Rodriguez-Bano J (2002) Nosocomial bacteremia due to Acinetobacter baumannii: epidemiology, clinical features and treatment. Clin Microbiol Infect 8: 687–693. doi: 10.1046/j.1469-0691.2002.00487.x
|
| [3] | Bonomo RA, Szabo D (2006) Mechanisms of multidrug resistance in Acinetobacter species and Pseudomonas aeruginosa. Clin Infect Dis 43 Suppl 2: S49–56. |
| [4] |
Devaud M, Kayser FH, Bächi B (1982) Transposon-mediated multiple antibiotic resistance in Acinetobacter strains. Antimicrob Agents Chemother 22: 323–329. doi: 10.1128/AAC.22.2.323
|
| [5] | Chu YW, Leung CM, Houang ET, et al. (1999) Skin carriage of Acinetobacters in Hong Kong. J Clin Microbiol 37: 2962–2967. |
| [6] |
Jawad A, Snelling AM, Heritage J, et al. (1998) Exceptional desiccation tolerance of Acinetobacter radioresistens. J Hosp Infect 39: 235–240. doi: 10.1016/S0195-6701(98)90263-8
|
| [7] |
Dijkshoorn L, Nemec A, Seifert H (2007) An increasing threat in hospitals: multidrug-resistant Acinetobacter baumannii. Nat Rev Microbiol 5: 939–951. doi: 10.1038/nrmicro1789
|
| [8] | Lopes JM, Goulart EM, Starling CE (2007) Pediatric mortality due to nosocomial infection: a critical approach. Braz J Infect Dis 11: 515–519. |
| [9] | Kurcik-Trajkovska B (2009) Acinetobacter spp. - A serious enemy threatening hospitals worldwide. Maced J Med Sci 2: 157–162. |
| [10] | La Scola B, Fournier PE, Brouqui P, et al. (2001) Detection and culture of Bartonella quintana, Serratia marcescens, and Acinetobacter spp. from decontaminated human body lice. J Clin Microbiol 39: 1707–1709. |
| [11] |
Tomaras AP, Dorsey CW, Edelmann RE, et al. (2003) Attachment to and biofilm formation on abiotic surfaces by Acinetobacter baumannii: involvement of a novel chaperone-usher pili assembly system. Microbiology 149: 3473–3484. doi: 10.1099/mic.0.26541-0
|
| [12] |
Lee JC, Koerten H, van den Broek P, et al. (2006) Adherence of Acinetobacter baumannii strains to human bronchial epithelial cells. Res Microbiol 157: 360–366. doi: 10.1016/j.resmic.2005.09.011
|
| [13] | Gospodarek E, Grzanka A, Dudziak Z, et al. (1998) Electron-microscopic observation of adherence of Acinetobacter baumannii to red blood cells. Acta Microbiol Pol 47: 213–217. |
| [14] |
Costerton JW, Stewart PS, Greenberg EP (1999) Bacterial biofilms: a common cause of persistent infections. Science 284: 1318–1322. doi: 10.1126/science.284.5418.1318
|
| [15] |
Rao RS, Karthika RU, Singh SP, et al. (2008) Correlation between biofilm production and multiple drug resistance in imipenem resistant clinical isolates of Acinetobacter baumannii. Indian J Med Microbiol 26: 333–337. doi: 10.4103/0255-0857.43566
|
| [16] |
Carpentier B, Cerf O (1993) Biofilms and their consequences, with particular reference to hygiene in the food industry. J Appl Bacteriol 75: 499–511. doi: 10.1111/j.1365-2672.1993.tb01587.x
|
| [17] |
Gilbert P, Brown MR (1998) Biofilms and beta-lactam activity. J Antimicrob Chemother 41: 571–572. doi: 10.1093/jac/41.5.571
|
| [18] | Hausner M, Wuertz S (1999) High rates of conjugation in bacterial biofilms as determined by quantitative in situ analysis. Appl Environ Microbiol 65: 3710–3713. |
| [19] |
Otto M (2009) Staphylococcus epidermidis--the 'accidental' pathogen. Nat Rev Microbiol 7: 555–567. doi: 10.1038/nrmicro2182
|
| [20] |
Peleg AY, Seifert H, Paterson DL (2008) Acinetobacter baumannii: emergence of a successful pathogen. Clin Microbiol Rev 21: 538–582. doi: 10.1128/CMR.00058-07
|
| [21] |
Choi CH, Lee EY, Lee YC, et al. (2005) Outer membrane protein 38 of Acinetobacter baumannii localizes to the mitochondria and induces apoptosis of epithelial cells. Cell Microbiol 7: 1127–1138. doi: 10.1111/j.1462-5822.2005.00538.x
|
| [22] |
Loehfelm TW, Luke NR, Campagnari AA (2008) Identification and characterization of an Acinetobacter baumannii biofilm-associated protein. J Bacteriol 190: 1036–1044. doi: 10.1128/JB.01416-07
|
| [23] |
Choi AH, Slamti L, Avci FY, et al. (2009) The pgaABCD locus of Acinetobacter baumannii encodes the production of poly-beta-1-6-N-acetylglucosamine, which is critical for biofilm formation. J Bacteriol 191: 5953–5963. doi: 10.1128/JB.00647-09
|
| [24] |
Gaddy JA, Tomaras AP, Actis LA (2009) The Acinetobacter baumannii 19606 OmpA protein plays a role in biofilm formation on abiotic surfaces and in the interaction of this pathogen with eukaryotic cells. Infect Immun 77: 3150–3160. doi: 10.1128/IAI.00096-09
|
| [25] |
Kim SW, Choi CH, Moon DC, et al. (2009) Serum resistance of Acinetobacter baumannii through the binding of factor H to outer membrane proteins. FEMS Microbiol Lett 301: 224–231. doi: 10.1111/j.1574-6968.2009.01820.x
|
| [26] |
Cabral MP, Soares NC, Aranda J, et al. (2011) Proteomic and functional analyses reveal a unique lifestyle for Acinetobacter baumannii biofilms and a key role for histidine metabolism. J Proteome Res 10: 3399–3417. doi: 10.1021/pr101299j
|
| [27] |
Choi CH, Lee JS, Lee YC, et al. (2008) Acinetobacter baumannii invades epithelial cells and outer membrane protein A mediates interactions with epithelial cells. BMC Microbiol 8: 216. doi: 10.1186/1471-2180-8-216
|
| [28] |
Clemmer KM, Bonomo RA, Rather PN (2011) Genetic analysis of surface motility in Acinetobacter baumannii. Microbiology 157: 2534–2544. doi: 10.1099/mic.0.049791-0
|
| [29] |
Itoh Y, Rice JD, Goller C, et al. (2008) Roles of pgaABCD genes in synthesis, modification, and export of the Escherichia coli biofilm adhesin poly-beta-1,6-N-acetyl-D-glucosamine. J Bacteriol 190: 3670–3680. doi: 10.1128/JB.01920-07
|
| [30] | Cramton SE, Gerke C, Schnell NF, et al. (1999) The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation. Infect Immun 67: 5427–5433. |
| [31] |
Kropec A, Maira-Litran T, Jefferson KK, et al. (2005) Poly-N-acetylglucosamine production in Staphylococcus aureus is essential for virulence in murine models of systemic infection. Infect Immun 73: 6868–6876. doi: 10.1128/IAI.73.10.6868-6876.2005
|
| [32] |
Lewis K (2001) Riddle of biofilm resistance. Antimicrob Agents Chemother 45: 999–1007. doi: 10.1128/AAC.45.4.999-1007.2001
|
| [33] |
Tomaras AP, Flagler MJ, Dorsey CW, et al. (2008) Characterization of a two-component regulatory system from Acinetobacter baumannii that controls biofilm formation and cellular morphology. Microbiology 154: 3398–3409. doi: 10.1099/mic.0.2008/019471-0
|
| [34] |
Luo LM, Wu LJ, Xiao YL, et al. (2015) Enhancing pili assembly and biofilm formation in Acinetobacter baumannii ATCC19606 using non-native acyl-homoserine lactones. BMC Microbiol 15: 62. doi: 10.1186/s12866-015-0397-5
|
| [35] |
Mussi MA, Limansky AS, Viale AM (2005) Acquisition of resistance to carbapenems in multidrug-resistant clinical strains of Acinetobacter baumannii: natural insertional inactivation of a gene encoding a member of a novel family of beta-barrel outer membrane proteins. Antimicrob Agents Chemother 49: 1432–1440. doi: 10.1128/AAC.49.4.1432-1440.2005
|
| [36] |
Poirel L, Lebessi E, Héritier C, et al. (2006) Nosocomial spread of OXA-58-positive carbapenem-resistant Acinetobacter baumannii isolates in a paediatric hospital in Greece. Clin Microbiol Infect 12: 1138–1141. doi: 10.1111/j.1469-0691.2006.01537.x
|
| [37] |
Aranda J, Bardina C, Beceiro A, et al. (2011) Acinetobacter baumannii RecA protein in repair of DNA damage, antimicrobial resistance, general stress response, and virulence. J Bacteriol 193: 3740–3747. doi: 10.1128/JB.00389-11
|
| [38] |
Diggle SP, Crusz SA, Camara M (2007) Quorum sensing. Curr Biol 17: R907–910. doi: 10.1016/j.cub.2007.08.045
|
| [39] |
Uroz S, Dessaux Y, Oger P (2009) Quorum sensing and quorum quenching: the yin and yang of bacterial communication. Chembiochem 10: 205–216. doi: 10.1002/cbic.200800521
|
| [40] |
Whitehead NA, Barnard AM, Slater H, et al. (2001) Quorum-sensing in Gram-negative bacteria. FEMS Microbiol Rev 25: 365–404. doi: 10.1111/j.1574-6976.2001.tb00583.x
|
| [41] |
Holden I, Swift I, Williams I (2000) New signal molecules on the quorum-sensing block. Trends Microbiol 8: 101–104; discussion 103–104. doi: 10.1016/S0966-842X(00)01718-2
|
| [42] | Irie Y, Parsek MR (2008) Quorum sensing and microbial biofilms. Curr Top Microbiol Immunol 322: 67–84. |
| [43] |
Williams P (2006) Quorum sensing. Int J Med Microbiol 296: 57–59. doi: 10.1016/j.ijmm.2006.01.034
|
| [44] | Schaefer AL, Hanzelka BL, Eberhard A, et al. (1996) Quorum sensing in Vibrio fischeri: probing autoinducer-LuxR interactions with autoinducer analogs. J Bacteriol 178: 2897–2901. |
| [45] |
González RH, Nusblat A, Nudel BC (2001) Detection and characterization of quorum sensing signal molecules in Acinetobacter strains. Microbiol Res 155: 271–277. doi: 10.1016/S0944-5013(01)80004-5
|
| [46] | González RH, Dijkshoorn L, Van den Barselaar M, et al. (2009) Quorum sensing signal profile of Acinetobacter strains from nosocomial and environmental sources. Rev Argent Microbiol 41: 73–78. |
| [47] | Prashanth K, Vasanth T, Saranathan R, et al. (2012) Antibiotic resistance, biofilms and quorum 487 sensing in Acinetobacter species, In: Antibiotic resistant bacteria - A coninuous challenge in the 488 new millennium, Dr. Marina Pana (Ed.); Croatia : InTech, 179–212. |
| [48] |
Niu C, Clemmer KM, Bonomo RA, et al. (2008) Isolation and characterization of an autoinducer synthase from Acinetobacter baumannii. J Bacteriol 190: 3386–3392. doi: 10.1128/JB.01929-07
|
| [49] |
Stevens AM, Dolan KM, Greenberg EP (1994) Synergistic binding of the Vibrio fischeri LuxR transcriptional activator domain and RNA polymerase to the lux promoter region. Proc Natl Acad Sci U S A 91: 12619–12623. doi: 10.1073/pnas.91.26.12619
|
| [50] |
Egland KA, Greenberg EP (2001) Quorum sensing in Vibrio fischeri: analysis of the LuxR DNA binding region by alanine-scanning mutagenesis. J Bacteriol 183: 382–386. doi: 10.1128/JB.183.1.382-386.2001
|
| [51] |
Latifi A, Winson MK, Foglino M, et al. (1995) Multiple homologues of LuxR and LuxI control expression of virulence determinants and secondary metabolites through quorum sensing in Pseudomonas aeruginosa PAO1. Mol Microbiol 17: 333–343. doi: 10.1111/j.1365-2958.1995.mmi_17020333.x
|
| [52] |
Gray KM, Garey JR (2001) The evolution of bacterial LuxI and LuxR quorum sensing regulators. Microbiology 147: 2379–2387. doi: 10.1099/00221287-147-8-2379
|
| [53] |
Bhargava N, Sharma P, Capalash N (2010) Quorum sensing in Acinetobacter: an emerging pathogen. Crit Rev Microbiol 36: 349-360. doi: 10.3109/1040841X.2010.512269
|
| [54] |
Surette MG, Miller MB, Bassler BL (1999) Quorum sensing in Escherichia coli, Salmonella typhimurium, and Vibrio harveyi: a new family of genes responsible for autoinducer production. Proc Natl Acad Sci U S A 96: 1639–1644. doi: 10.1073/pnas.96.4.1639
|
| [55] |
Young DM, Parke D, Ornston LN (2005) Opportunities for genetic investigation afforded by Acinetobacter baylyi, a nutritionally versatile bacterial species that is highly competent for natural transformation. Annu Rev Microbiol 59: 519–551. doi: 10.1146/annurev.micro.59.051905.105823
|
| [56] |
Smith MG, Gianoulis TA, Pukatzki S, et al. (2007) New insights into Acinetobacter baumannii pathogenesis revealed by high-density pyrosequencing and transposon mutagenesis. Genes Dev 21: 601–614. doi: 10.1101/gad.1510307
|
| [57] |
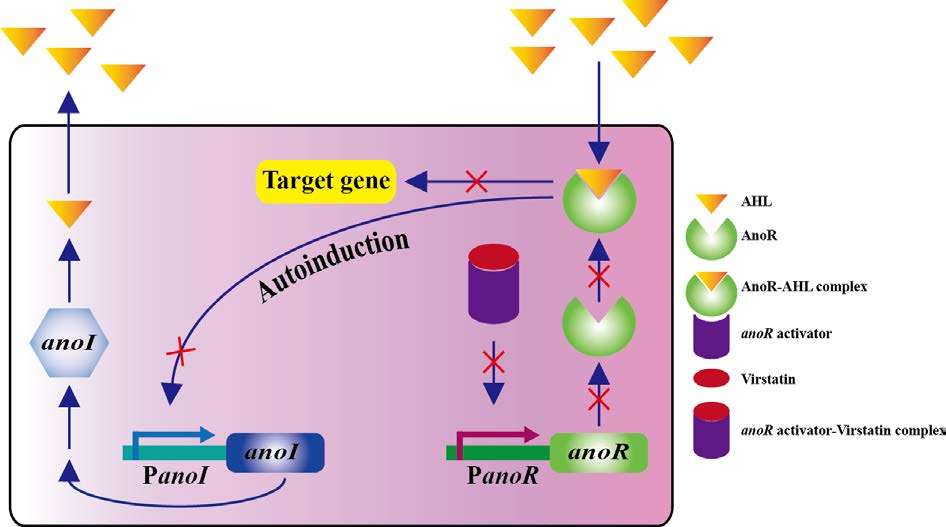
Oh MH, Choi CH (2015) Role of LuxIR homologue AnoIR in Acinetobacter nosocomialis and the effect of virstatin on the expression of anoR Gene. J Microbiol Biotechnol 25: 1390–1400. doi: 10.4014/jmb.1504.04069
|
| [58] |
Taccone FS, Rodriguez-Villalobos H, De Backer D, et al. (2006) Successful treatment of septic shock due to pan-resistant Acinetobacter baumannii using combined antimicrobial therapy including tigecycline. Eur J Clin Microbiol Infect Dis 25: 257–260. doi: 10.1007/s10096-006-0123-1
|
| [59] |
Valencia R, Arroyo LA, Conde M, et al. (2009) Nosocomial outbreak of infection with pan-drug-resistant Acinetobacter baumannii in a tertiary care university hospital. Infect Control Hosp Epidemiol 30: 257–263. doi: 10.1086/595977
|
| [60] | Amaral L, Martins A, Spengler G, et al. (2014) Efflux pumps of Gram-negative bacteria: what they do, how they do it, with what and how to deal with them. Front Pharmacol 4: 168. |
| [61] |
Balaban N, Cirioni O, Giacometti A, et al. (2007) Treatment of Staphylococcus aureus biofilm infection by the quorum-sensing inhibitor RIP. Antimicrob Agents Chemother 51: 2226–2229. doi: 10.1128/AAC.01097-06
|
| [62] |
Hoffman LR, D'Argenio DA, MacCoss MJ, et al. (2005) Aminoglycoside antibiotics induce bacterial biofilm formation. Nature 436: 1171–1175. doi: 10.1038/nature03912
|
| [63] |
Henikoff S, Wallace JC, Brown JP (1990) Finding protein similarities with nucleotide sequence databases. Methods Enzymol 183: 111–132. doi: 10.1016/0076-6879(90)83009-X
|
| [64] |
Rahmati S, Yang S, Davidson AL, et al. (2002) Control of the AcrAB multidrug efflux pump by quorum-sensing regulator SdiA. Mol Microbiol 43: 677–685. doi: 10.1046/j.1365-2958.2002.02773.x
|
| [65] |
Maseda H, Sawada I, Saito K, et al. (2004) Enhancement of the mexAB-oprM efflux pump expression by a quorum-sensing autoinducer and its cancellation by a regulator, MexT, of the mexEF-oprN efflux pump operon in Pseudomonas aeruginosa. Antimicrob Agents Chemother 48: 1320–1328. doi: 10.1128/AAC.48.4.1320-1328.2004
|
| [66] |
Chu YW, Chau SL, Houang ET (2006) Presence of active efflux systems AdeABC, AdeDE and AdeXYZ in different Acinetobacter genomic DNA groups. J Med Microbiol 55: 477–478. doi: 10.1099/jmm.0.46433-0
|
| [67] |
Nemec A, Maixnerova M, van der Reijden TJ, et al. (2007) Relationship between the AdeABC efflux system gene content, netilmicin susceptibility and multidrug resistance in a genotypically diverse collection of Acinetobacter baumannii strains. J Antimicrob Chemother 60: 483–489. doi: 10.1093/jac/dkm231
|
| [68] |
Yoon EJ, Courvalin P, Grillot-Courvalin C (2013) RND-type efflux pumps in multidrug-resistant clinical isolates of Acinetobacter baumannii: major role for AdeABC overexpression and AdeRS mutations. Antimicrob Agents Chemother 57: 2989–2995. doi: 10.1128/AAC.02556-12
|
| [69] |
Coyne S, Courvalin P, Périchon B (2011) Efflux-mediated antibiotic resistance in Acinetobacter spp. Antimicrob Agents Chemother 55: 947–953. doi: 10.1128/AAC.01388-10
|
| [70] |
He X, Lu F, Yuan F, et al. (2015) Biofilm formation caused by clinical Acinetobacter baumannii isolates is associated with overexpression of the AdeFGH Efflux pump. Antimicrob Agents Chemother 59: 4817–4825. doi: 10.1128/AAC.00877-15
|
| [71] |
Kayama S, Murakami K, Ono T, et al. (2009) The role of rpoS gene and quorum-sensing system in ofloxacin tolerance in Pseudomonas aeruginosa. FEMS Microbiol Lett 298: 184–192. doi: 10.1111/j.1574-6968.2009.01717.x
|
| [72] |
Que YA, Hazan R, Strobel B, et al. (2013) A quorum sensing small volatile molecule promotes antibiotic tolerance in bacteria. PLoS One 8: e80140. doi: 10.1371/journal.pone.0080140
|
| [73] |
Khan MS, Zahin M, Hasan S, et al. (2009) Inhibition of quorum sensing regulated bacterial functions by plant essential oils with special reference to clove oil. Lett Appl Microbiol 49: 354–360. doi: 10.1111/j.1472-765X.2009.02666.x
|
| [74] |
Sperandio V (2007) Novel approaches to bacterial infection therapy by interfering with bacteria-to-bacteria signaling. Expert Rev Anti Infect Ther 5: 271–276. doi: 10.1586/14787210.5.2.271
|
| [75] |
Stacy DM, Welsh MA, Rather PN, et al. (2012) Attenuation of quorum sensing in the pathogen Acinetobacter baumannii using non-native N-Acyl homoserine lactones. ACS Chem Biol 7: 1719–1728. doi: 10.1021/cb300351x
|
| [76] |
Saroj SD, Rather PN (2013) Streptomycin inhibits quorum sensing in Acinetobacter baumannii. Antimicrob Agents Chemother 57: 1926–1929. doi: 10.1128/AAC.02161-12
|
| [77] |
Chabane YN, Mlouka MB, Alexandre S, et al. (2014) Virstatin inhibits biofilm formation and motility of Acinetobacter baumannii. BMC Microbiol 14: 62. doi: 10.1186/1471-2180-14-62
|
| [78] | Choo JH, Rukayadi Y, Hwang JK (2006) Inhibition of bacterial quorum sensing by vanilla extract. Lett Appl Microbiol 42: 637–641. |
| [79] |
Rasmussen TB, Bjarnsholt T, Skindersoe ME, et al. (2005) Screening for quorum-sensing inhibitors (QSI) by use of a novel genetic system, the QSI selector. J Bacteriol 187: 1799–1814. doi: 10.1128/JB.187.5.1799-1814.2005
|
| [80] |
Cady NC, McKean KA, Behnke J, et al. (2012) Inhibition of biofilm formation, quorum sensing and infection in Pseudomonas aeruginosa by natural products-inspired organosulfur compounds. PLoS One 7: e38492. doi: 10.1371/journal.pone.0038492
|
| [81] |
Sambanthamoorthy K, Luo C, Pattabiraman N, et al. (2014) Identification of small molecules inhibiting diguanylate cyclases to control bacterial biofilm development. Biofouling 30: 17–28. doi: 10.1080/08927014.2013.832224
|
| [82] |
Dong YH, Wang LH, Xu JL, et al. (2001) Quenching quorum-sensing-dependent bacterial infection by an N-acyl homoserine lactonase. Nature 411: 813–817. doi: 10.1038/35081101
|
| [83] |
Chow JY, Yang Y, Tay SB, et al. (2014) Disruption of biofilm formation by the human pathogen Acinetobacter baumannii using engineered quorum-quenching lactonases. Antimicrob Agents Chemother 58: 1802–1805. doi: 10.1128/AAC.02410-13
|
| [84] | Hoang TT, Schweizer HP (1999) Characterization of Pseudomonas aeruginosa enoyl-acyl carrier protein reductase (FabI): a target for the antimicrobial triclosan and its role in acylated homoserine lactone synthesis. J Bacteriol 181: 5489–5497. |
| [85] |
Jarrett CO, Deak E, Isherwood KE, et al. (2004) Transmission of Yersinia pestis from an infectious biofilm in the flea vector. J Infect Dis 190: 783–792. doi: 10.1086/422695
|
| [86] | Czajkowski R, Jafra S (2009) Quenching of acyl-homoserine lactone-dependent quorum sensing by enzymatic disruption of signal molecules. Acta Biochim Pol 56: 1–16. |
| [87] |
Lin YH, Xu JL, Hu J, et al. (2003) Acyl-homoserine lactone acylase from Ralstonia strain XJ12B represents a novel and potent class of quorum-quenching enzymes. Mol Microbiol 47: 849–860. doi: 10.1046/j.1365-2958.2003.03351.x
|
| [88] | Romero M, Diggle SP, Heeb S, et al. (2008) Quorum quenching activity in Anabaena sp. PCC 7120: identification of AiiC, a novel AHL-acylase. FEMS Microbiol Lett 280: 73–80. |
| [89] |
Uroz S, Oger PM, Chapelle E, et al. (2008) A Rhodococcus qsdA-encoded enzyme defines a novel class of large-spectrum quorum-quenching lactonases. Appl Environ Microbiol 74: 1357–1366. doi: 10.1128/AEM.02014-07
|
| [90] |
Chow JY, Xue B, Lee KH, et al. (2010) Directed evolution of a thermostable quorum-quenching lactonase from the amidohydrolase superfamily. J Biol Chem 285: 40911–40920. doi: 10.1074/jbc.M110.177139
|
| [91] | Kiran S (2011) Enzymatic quorum quenching increases antibiotic susceptibility of multidrug resistant Pseudomonas aeruginosa. Irani J Microbiol 3: 1–12. |
| [92] |
Limsuwan S, Subhadhirasakul S, Voravuthikunchai PS (2009) Medicinal plants with significant activity against important pathogenic bacteria. Pharm Biol 47: 683–689. doi: 10.1080/13880200902930415
|
| [93] |
Babić F, Venturi V, Maravić-Vlahoviček G (2010) Tobramycin at subinhibitory concentration inhibits the RhlI/R quorum sensing system in a Pseudomonas aeruginosa environmental isolate. BMC Infect Dis 10: 148. doi: 10.1186/1471-2334-10-148
|
| [94] |
Defoirdt T, Boon N, Bossier P (2010) Can bacteria evolve resistance to quorum sensing disruption? PLoS Pathog 6: e1000989. doi: 10.1371/journal.ppat.1000989
|
Figures(1)

Bindu Subhadra, Man Hwan Oh, Chul Hee Choi. Quorum sensing in Acinetobacter: with special emphasis on antibiotic resistance, biofilm formation and quorum quenching[J]. AIMS Microbiology, 2016, 2(1): 27-41. doi: 10.3934/microbiol.2016.1.27







 DownLoad:
DownLoad: