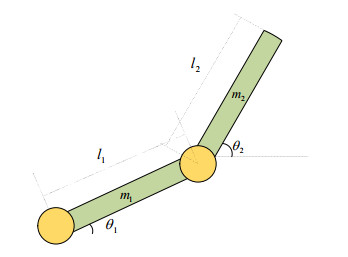
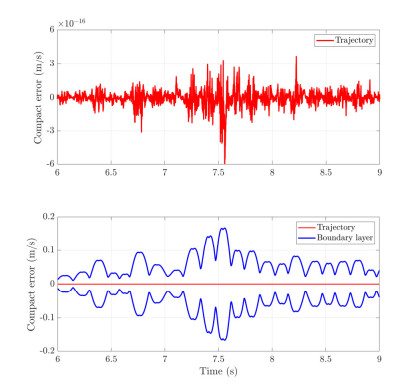
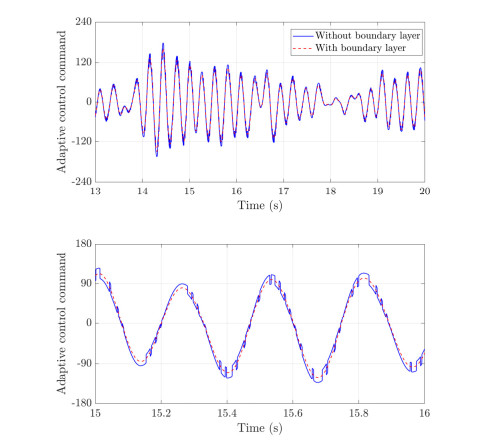
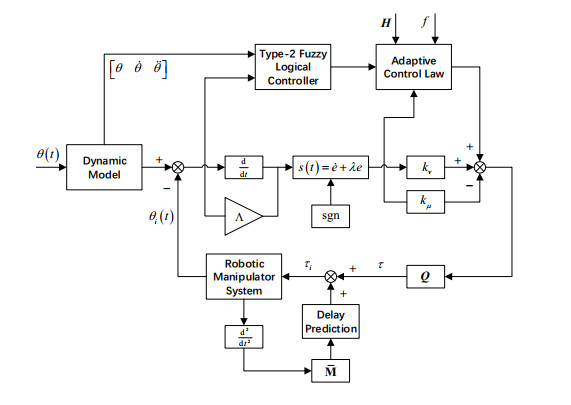
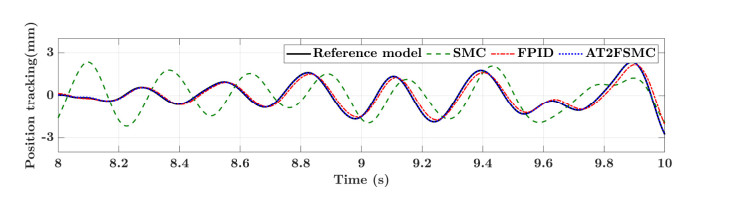
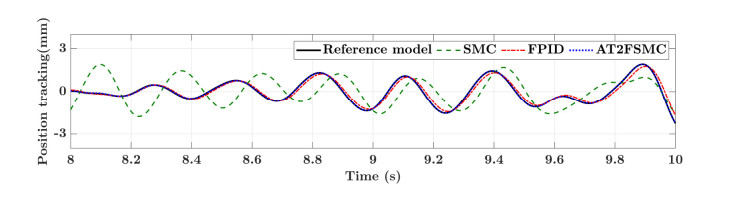
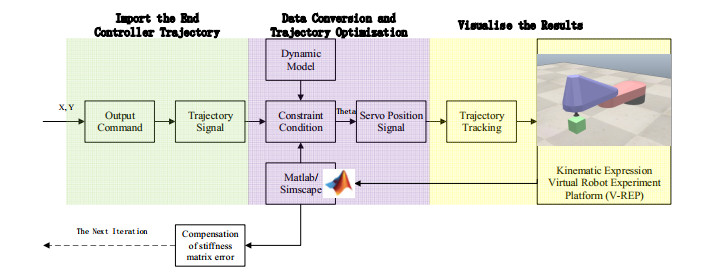
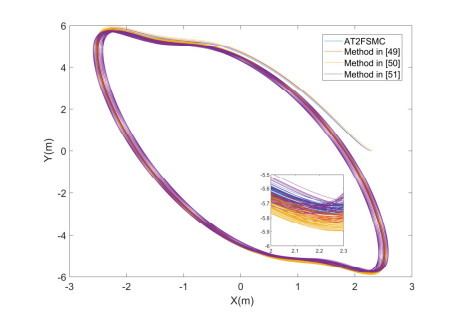
With the wide application of intelligent manufacturing and the development of diversified functions of industrial manipulator, the requirements for the control accuracy and stability of the manipulator servo system are also increasing. The control of industrial manipulator is a time-varying system with nonlinear and strong coupling, which is often affected by uncertain factors, including parameter changing, environmental interference, joint friction and so on. Aiming at the problem of the poor control accuracy of the manipulator. Under the complex disturbance environment, control accuracy of the manipulator will be greatly affected, so this paper proposes an adaptive type-2 fuzzy sliding mode control (AT2FSMC) method applied to the servo control of the industrial manipulator, which realizes the adaptive adjustment of the boundary layer thickness to suppress the trajectory error caused by the external disturbance and weakens the chattering problem of the sliding mode control. The simulation results on a two-axis manipulator indicate that, with the presence of external disturbances, the proposed control method can help the manipulator maintain control signal stability and improve tracking accuracy. It also suppressed chattering produced by sliding mode control (SMC) and strengthening the robustness of the system. Compared with other conventional trajectory tracking control methods, the effectiveness of the proposed method can be reflected. Finally, the proposed method is tested in an actual manipulator to complete a practical trajectory to prove its feasibility.
Citation: Qianqian Zhang, Mingye Mu, Heyuan Ji, Qiushi Wang, Xingyu Wang. An adaptive type-2 fuzzy sliding mode tracking controller for a robotic manipulator[J]. Electronic Research Archive, 2023, 31(7): 3791-3813. doi: 10.3934/era.2023193
With the wide application of intelligent manufacturing and the development of diversified functions of industrial manipulator, the requirements for the control accuracy and stability of the manipulator servo system are also increasing. The control of industrial manipulator is a time-varying system with nonlinear and strong coupling, which is often affected by uncertain factors, including parameter changing, environmental interference, joint friction and so on. Aiming at the problem of the poor control accuracy of the manipulator. Under the complex disturbance environment, control accuracy of the manipulator will be greatly affected, so this paper proposes an adaptive type-2 fuzzy sliding mode control (AT2FSMC) method applied to the servo control of the industrial manipulator, which realizes the adaptive adjustment of the boundary layer thickness to suppress the trajectory error caused by the external disturbance and weakens the chattering problem of the sliding mode control. The simulation results on a two-axis manipulator indicate that, with the presence of external disturbances, the proposed control method can help the manipulator maintain control signal stability and improve tracking accuracy. It also suppressed chattering produced by sliding mode control (SMC) and strengthening the robustness of the system. Compared with other conventional trajectory tracking control methods, the effectiveness of the proposed method can be reflected. Finally, the proposed method is tested in an actual manipulator to complete a practical trajectory to prove its feasibility.

| [1] |
E. Tatlicioglu, D. Braganza, T. C. Burg, D. M. Dawson, Adaptive control of redundant robot manipulators with sub-task objectives, Robotica, 27 (2009), 873–881. https://doi.org/10.1017/S0263574708005274 doi: 10.1017/S0263574708005274

|
| [2] |
M. D. Tran, H. J. Kang, Adaptive terminal sliding mode control of uncertain robotic manipulators based on local approximation of a dynamic system, Neurocomputing, 228 (2017), 231–240. https://doi.org/10.1016/j.neucom.2016.09.089 doi: 10.1016/j.neucom.2016.09.089
|
| [3] |
R. G. Roy, D. Ghoshal, Grey wolf optimization-based second order sliding mode control for inchworm robot, Robotica, 38 (2020), 1539–1557. https://doi.org/10.1017/S0263574719001620 doi: 10.1017/S0263574719001620
|
| [4] |
A. Azizi, Applications of artificial intelligence techniques to enhance sustainability of industry 4.0: design of an artificial neural network model as dynamic behavior optimizer of robotic arms, Complexity, 2020 (2020), 8564140. https://doi.org/10.1155/2020/8564140 doi: 10.1155/2020/8564140
|
| [5] |
L. Xie, X. Yu, L. Chen, Robust fuzzy sliding mode control and vibration suppression of free-floating flexible-link and flexible-joints space manipulator with external interference and uncertain parameter, Robotica, 40 (2022), 997–1019. https://doi.org/10.1017/S0263574721000977 doi: 10.1017/S0263574721000977
|
| [6] |
S. Sui, T. Zhao, Active disturbance rejection control for optoelectronic stabilized platform based on adaptive fuzzy sliding mode control, ISA Trans., 125 (2022), 85–98. https://doi.org/10.1016/j.isatra.2021.06.020 doi: 10.1016/j.isatra.2021.06.020
|
| [7] |
A. Rosales, L. Ibarra, P. Ponce, A. Molina, Fuzzy sliding mode control design based on stability margins, J. Franklin Inst., 356 (2019), 5260–5273. https://doi.org/10.1016/j.jfranklin.2019.04.035 doi: 10.1016/j.jfranklin.2019.04.035
|
| [8] |
S. Mobayen, J. Ma, Robust finite-time composite nonlinear feedback control for synchronization of uncertain chaotic systems with nonlinearity and time-delay, Chaos, Solitons Fractals, 114 (2018), 46–54. https://doi.org/10.1016/j.chaos.2018.06.020 doi: 10.1016/j.chaos.2018.06.020
|
| [9] |
H. Feng, Q. Song, S. Ma, W. Ma, C. Yin, D. Cao, et al., A new adaptive sliding mode controller based on the RBF neural network for an electro-hydraulic servo system, ISA Trans., 129 (2022), 472–484. https://doi.org/10.1016/j.isatra.2021.12.044 doi: 10.1016/j.isatra.2021.12.044
|
| [10] |
K. Shao, J. Zheng, R. Tang, X. Li, Z. Man, B. Liang, Barrier function based adaptive sliding mode control for uncertain systems with input saturation, IEEE/ASME Trans. Mechatron., 27 (2022), 4258–4268. https://doi.org/10.1109/TMECH.2022.3153670 doi: 10.1109/TMECH.2022.3153670
|
| [11] |
P. Shi, M. Liu, L. Zhang, Fault-tolerant sliding-mode-observer synthesis of Markovian jump systems using quantized measurements, IEEE Trans. Ind. Electron., 62 (2015), 5910–5918. https://doi.org/10.1109/TIE.2015.2442221 doi: 10.1109/TIE.2015.2442221
|
| [12] |
M. Van, Higher-order terminal sliding mode controller for fault accommodation of Lipschitz second-order nonlinear systems using fuzzy neural network, Appl. Soft Comput., 104 (2021), 107186. https://doi.org/10.1016/j.asoc.2021.107186 doi: 10.1016/j.asoc.2021.107186
|
| [13] |
S. Mobayen, F. Tchier, Nonsingular fast terminal sliding-mode stabilizer for a class of uncertain nonlinear systems based on disturbance observer, Sci. Iran., 24 (2017), 1410–1418. https://doi.org/10.24200/sci.2017.4123 doi: 10.24200/sci.2017.4123
|
| [14] |
S. Ding, J. Wang, W. X. Zheng, Second-order sliding mode control for nonlinear uncertain systems bounded by positive functions, IEEE Trans. Ind. Electron., 62 (2015), 5899–5909. https://doi.org/10.1109/TIE.2015.2448064 doi: 10.1109/TIE.2015.2448064
|
| [15] |
S. Laghrouche, F. Plestan, A. Glumineau, Higher order sliding mode control based on integral sliding mode, Automatica, 43 (2007), 531–537. https://doi.org/10.1016/j.automatica.2006.09.017 doi: 10.1016/j.automatica.2006.09.017
|
| [16] |
S. Li, M. Zhou, X. Yu, Design and implementation of terminal sliding mode control method for PMSM speed regulation system, IEEE Trans. Ind. Inf., 9 (2013), 1879–1891. https://doi.org/10.1109/TⅡ.2012.2226896 doi: 10.1109/TⅡ.2012.2226896
|
| [17] |
S. Mobayen, Design of LMI-based sliding mode controller with an exponential policy for a class of underactuated systems, Complexity, 21 (2016), 117–124. https://doi.org/10.1002/cplx.21636 doi: 10.1002/cplx.21636
|
| [18] |
J. Liu, S. Laghrouche, M. Harmouche, M. Wack, Adaptive-gain second-order sliding mode observer design for switching power converters, Control Eng. Pract., 30 (2014), 124–131. https://doi.org/10.1016/j.conengprac.2013.10.012 doi: 10.1016/j.conengprac.2013.10.012
|
| [19] |
J. Liu, S. Laghrouche, M. Wack, Observer-based higher order sliding mode control of power factor in three-phase AC/DC converter for hybrid electric vehicle applications, Int. J. Control, 87 (2014), 1117–1130. https://doi.org/10.1080/00207179.2013.868609 doi: 10.1080/00207179.2013.868609
|
| [20] |
L. Bao, D. Kim, S. Yi, J. Lee, Design of a sliding mode controller with fuzzy rules for a 4-DoF service robot, Int. J. Control Autom. Syst., 19 (2021), 2869–2881. https://doi.org/10.1007/s12555-020-0452-3 doi: 10.1007/s12555-020-0452-3
|
| [21] |
L. Wang, T. Chai, L. Zhai, Neural-network-based terminal sliding-mode control of robotic manipulators including actuator dynamics, IEEE Trans. Ind. Electron., 56 (2009), 3296–3304. https://doi.org/10.1109/TIE.2008.2011350 doi: 10.1109/TIE.2008.2011350
|
| [22] |
T. Sun, H. Pei, Y. Pan, H. Zhou, C. Zhang, Neural network-based sliding mode adaptive control for robot manipulators, Neurocomputing, 74 (2011), 2377–2384. https://doi.org/10.1016/j.neucom.2011.03.015 doi: 10.1016/j.neucom.2011.03.015
|
| [23] |
J. Baek, M. Jin, S. Han, A new adaptive sliding mode control scheme for application to robot manipulators, IEEE Trans. Ind. Electron., 63 (2016), 3628–3637. https://doi.org/10.1109/TIE.2016.2522386 doi: 10.1109/TIE.2016.2522386
|
| [24] |
Y. Su, Comments on "A new adaptive sliding-mode control scheme for application to robot manipulators", IEEE Trans. Ind. Electron., 67 (2020), 7116–7120. https://doi.org/10.1109/TIE.2019.2939995 doi: 10.1109/TIE.2019.2939995
|
| [25] |
A. Al-Jodah, B. Shirinzadeh, M. Ghafarian, T. K. Das, Y. Tian, D. Zhang, A fuzzy disturbance observer based control approach for a novel 1-DOF micropositioning mechanism, Mechatronics, 65 (2020), 102317. https://doi.org/10.1016/j.mechatronics.2019.102317 doi: 10.1016/j.mechatronics.2019.102317
|
| [26] |
M. Ghafarian, B. Shirinzadeh, A. Al-Jodah, T. K. Das, Adaptive fuzzy sliding mode control for high-precision motion tracking of a multi-DOF micro/nano manipulator, IEEE Rob. Autom. Lett., 5 (2020), 4313–4320. https://doi.org/10.1109/LRA.2020.2996065 doi: 10.1109/LRA.2020.2996065
|
| [27] |
J. Lee, P. H. Chang, M. Jin, Adaptive integral sliding mode control with time-delay estimation for robot manipulators, IEEE Trans. Ind. Electron., 64 (2017), 6796–6804. https://doi.org/10.1109/TIE.2017.2698416 doi: 10.1109/TIE.2017.2698416
|
| [28] |
B. Xiao, S. Yin, O. Kaynak, Tracking control of robotic manipulators with uncertain kinematics and dynamics, IEEE Trans. Ind. Electron., 63 (2016), 6439–6449. https://doi.org/10.1109/TIE.2016.2569068 doi: 10.1109/TIE.2016.2569068
|
| [29] |
A. A. L. Al-Jodah, B. Shirinzadeh, J. Pinskier, M. Ghafarian, T. K. Das, Y. Tian, et al., Antlion optimized robust control approach for micropositioning trajectory tracking tasks, IEEE Access, 8 (2020), 220889–220907. https://doi.org/10.1109/ACCESS.2020.3043411 doi: 10.1109/ACCESS.2020.3043411
|
| [30] |
M. Latifinavid, A. Azizi, Development of a vision-based unmanned ground vehicle for mapping and tennis ball collection: a fuzzy logic approach, Future Internet, 15 (2023), 84. https://doi.org/10.3390/fi15020084 doi: 10.3390/fi15020084
|
| [31] |
M. Latifinavid, A. Azizi, Kinematic modelling and position control of a 3-DOF parallel stabilizing robot manipulator, J. Intell. Rob. Syst., 107 (2023), 17. https://doi.org/10.1007/s10846-022-01795-x doi: 10.1007/s10846-022-01795-x
|
| [32] |
A. Li, M. Liu, X. Cao, R. Liu, Adaptive quantized sliding mode attitude tracking control for flexible spacecraft with input dead-zone via Takagi-Sugeno fuzzy approach, Inf. Sci., 587 (2022), 746–773. https://doi.org/10.1016/j.ins.2021.11.002 doi: 10.1016/j.ins.2021.11.002
|
| [33] |
S. Wen, T. Huang, X. Yu, M. Z. Q. Chen, Z. Zeng, Aperiodic sampled-data sliding-mode control of fuzzy systems with communication delays via the event-triggered method, IEEE Trans. Fuzzy Syst., 24 (2016), 1048–1057. https://doi.org/10.1109/TFUZZ.2015.2501412 doi: 10.1109/TFUZZ.2015.2501412
|
| [34] |
H. Liu, T. Zhang, Fuzzy sliding mode control of robotic manipulators with kinematic and dynamic uncertainties, J. Dyn. Syst. Meas. Contr., 134 (2012), 061007. https://doi.org/10.1115/1.4006626 doi: 10.1115/1.4006626
|
| [35] |
Z. Zhu, Y. Xia, M. Fu, Adaptive sliding mode control for attitude stabilization with actuator saturation, IEEE Trans. Ind. Electron., 58 (2011), 4898–4907. https://doi.org/10.1109/TIE.2011.2107719 doi: 10.1109/TIE.2011.2107719
|
| [36] |
N. T. T. Vu, D. Y. Yu, H. H. Choi, J. W. Jung, T–S fuzzy-model-based sliding-mode control for surface-mounted permanent-magnet synchronous motors considering uncertainties, IEEE Trans. Ind. Electron., 60 (2013), 4281–4291. https://doi.org/10.1109/TIE.2012.2213554 doi: 10.1109/TIE.2012.2213554
|
| [37] |
L. Qu, W. Qiao, L. Qu, Active-disturbance-rejection-based sliding-mode current control for permanent-magnet synchronous motors, IEEE Trans. Ind. Electron., 36 (2020), 751–760. https://doi.org/10.1109/TPEL.2020.3003666 doi: 10.1109/TPEL.2020.3003666
|
| [38] |
N. Nasiri, A. Fakharian, M. B. Menhaj, Observer-based robust control for flexible-joint robot manipulators: a state-dependent Riccati equation-based approach, Trans. Inst. Meas. Control, 42 (2020), 3135–3155. https://doi.org/10.1177/0142331220941653 doi: 10.1177/0142331220941653
|
| [39] |
I. H. Li, H. H. Chiang, L.W. Lee, Development of a linear delta robot with three horizontal-axial pneumatic actuators for 3-DOF trajectory tracking, Appl. Sci., 10 (2020), 3526. https://doi.org/10.3390/app10103526 doi: 10.3390/app10103526
|
| [40] |
I. Castillo, L. B. Freidovich, Describing-function-based analysis to tune parameters of chattering reducing approximations of sliding mode controllers, Control Eng. Pract., 95 (2020), 104230. https://doi.org/10.1016/j.conengprac.2019.104230 doi: 10.1016/j.conengprac.2019.104230
|
| [41] |
S. K. Wan, X. H. Li, W. J. Su, J. P. Yuan, J. Hong, Active chatter suppression for milling process with sliding mode control and electromagnetic actuator, Mech. Syst. Signal Process., 136 (2020), 106528. https://doi.org/10.1016/j.ymssp.2019.106528 doi: 10.1016/j.ymssp.2019.106528
|
| [42] | S. Kim, S. Choi, H. J. Kim, Aerial manipulation using a quadrotor with a two DOF robotic arm, in 2013 IEEE/RSJ International Conference on Intelligent Robots and Systems, (2013), 4990–4995. https://doi.org/10.1109/IROS.2013.6697077 |
| [43] | M. Sadeghijaleh, Voltage control strategy for direct-drive robots driven by permanent magnet synchronous motors, Int. J. Eng. Trans. B, 28 (2015), 709–716. Available from: https://www.ije.ir/article_72509.html. |
| [44] |
G. S. Lima, S. Trimpe, W. M. Bessa, Sliding mode control with gaussian process regression for underwater robots, J. Intell. Rob. Syst., 99 (2020), 487–498. https://doi.org/10.1007/s10846-019-01128-5 doi: 10.1007/s10846-019-01128-5
|
| [45] |
T. Wang, W. F. Xie, Y. M. Zhang, Sliding mode fault tolerant control dealing with modeling uncertainties and actuator faults, ISA Trans., 51 (2012), 386–392. https://doi.org/10.1016/j.isatra.2012.02.003 doi: 10.1016/j.isatra.2012.02.003
|
| [46] |
Z. H. Mao, X. G. Yan, B. Jiang, M. Chen, Adaptive fault-tolerant sliding-mode control for high-speed trains with actuator faults and uncertainties, IEEE Trans. Intell. Transp. Syst., 21 (2020), 2449–2460. https://doi.org/10.1109/TITS.2019.2918543 doi: 10.1109/TITS.2019.2918543
|
| [47] |
Q. L. Hu, B. Xiao, Adaptive fault tolerant control using integral sliding mode strategy with application to flexible spacecraft, Int. J. Syst. Sci., 44 (2013), 2273–2286. https://doi.org/10.1080/00207721.2012.702236 doi: 10.1080/00207721.2012.702236
|
| [48] | D. Wu, J. M. Mendel, Designing practical interval type-2 fuzzy logic systems made simple, in 2014 IEEE International Conference on Fuzzy Systems (FUZZ-IEEE), (2014), 800–807. https://doi.org/10.1109/FUZZ-IEEE.2014.6891534 |
| [49] |
R. R. Ardeshiri, M. H. Khooban, A. Noshadi, N. Vafamand, M. Rakhshan, Robotic manipulator control based on an optimal fractional-order fuzzy PID approach: SiL real-time simulation, Soft Comput., 24 (2020), 3849–3860. https://doi.org/10.1007/s00500-019-04152-7 doi: 10.1007/s00500-019-04152-7
|
| [50] |
D. Li, H. Yu, K. P. Tee, Y. Wu, S. S. Ge, T. H. Lee, On time-synchronized stability and control, IEEE Trans. Syst. Man Cybern.: Syst., 52 (2021), 2450–2463. https://doi.org/10.1109/TSMC.2021.3050183 doi: 10.1109/TSMC.2021.3050183
|
| [51] |
D. Li, S. S. Ge, T. H. Lee, Simultaneous arrival to origin convergence: sliding-mode control through the norm-normalized sign function, IEEE Trans. Autom. Control, 67 (2021), 1966–1972. https://doi.org/10.1109/TAC.2021.3069816 doi: 10.1109/TAC.2021.3069816
|



Figures(15) / Tables(4)
Qianqian Zhang, Mingye Mu, Heyuan Ji, Qiushi Wang, Xingyu Wang. An adaptive type-2 fuzzy sliding mode tracking controller for a robotic manipulator[J]. Electronic Research Archive, 2023, 31(7): 3791-3813. doi: 10.3934/era.2023193







 DownLoad:
DownLoad: