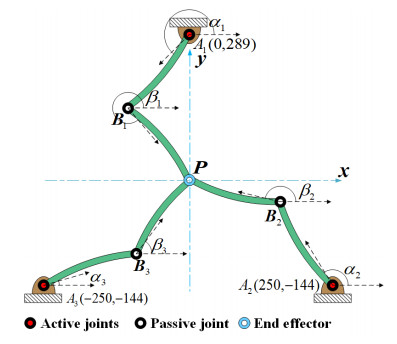
The application of a high-speed parallel manipulator necessitates the adoption of a lightweight design to reduce dead weight. However, this increases the elastic deformation of certain components, affecting the dynamic performance of the system. This study examined a 2-DOF planar flexible parallel manipulator. A dynamic model of the parallel manipulator composed of fully flexible links was established using a floating reference coordinate system and a combination of the finite element and augmented Lagrange multiplier methods. A dynamic analysis of the simplified model under three driving torque modes showed that the axial deformation was less than the transverse deformation by three orders of magnitude. Further, the kinematic and dynamic performance of the redundant drive was significantly better than that of the non-redundant drive, and the vibration was well suppressed in the redundant drive mode. In addition, the comprehensive performance of driving Mode 2 was better than that of the other two modes. Finally, the validity of the dynamic model was verified by modeling via Adams. The modular modeling method is conducive to the extension to other models and programming. Furthermore, the dynamic model of the established fully flexible link system can aid in optimizing the lightweight design and dynamic performance of the parallel manipulator.
Citation: Zhen Liu, Song Yang, Chen Cheng, Tao Ding, Ruimin Chai. Study on modeling and dynamic performance of a planar flexible parallel manipulator based on finite element method[J]. Mathematical Biosciences and Engineering, 2023, 20(1): 807-836. doi: 10.3934/mbe.2023037
The application of a high-speed parallel manipulator necessitates the adoption of a lightweight design to reduce dead weight. However, this increases the elastic deformation of certain components, affecting the dynamic performance of the system. This study examined a 2-DOF planar flexible parallel manipulator. A dynamic model of the parallel manipulator composed of fully flexible links was established using a floating reference coordinate system and a combination of the finite element and augmented Lagrange multiplier methods. A dynamic analysis of the simplified model under three driving torque modes showed that the axial deformation was less than the transverse deformation by three orders of magnitude. Further, the kinematic and dynamic performance of the redundant drive was significantly better than that of the non-redundant drive, and the vibration was well suppressed in the redundant drive mode. In addition, the comprehensive performance of driving Mode 2 was better than that of the other two modes. Finally, the validity of the dynamic model was verified by modeling via Adams. The modular modeling method is conducive to the extension to other models and programming. Furthermore, the dynamic model of the established fully flexible link system can aid in optimizing the lightweight design and dynamic performance of the parallel manipulator.

| [1] |
C. Gosselin, J. Angeles, Singularity analysis of closed-loop kinematic chains, IEEE Trans. Rob. Automat, 6 (1990), 281–290. https://doi.org/10.1109/70.56660 doi: 10.1109/70.56660

|
| [2] |
H. Ye, D. Wang, J. Wu, Y. Yue, Y. Zhou, Forward and inverse kinematics of a 5-DOF hybrid robot for composite material machining, Rob. Comput. Integr. Manuf., 65 (2020), 101961. https://doi.org/10.1016/j.rcim.2020.101961 doi: 10.1016/j.rcim.2020.101961
|
| [3] |
K. Dong, D. Li, X. Xue, C. Xu, H. Wang, X. Gao, Workspace and accuracy analysis on a novel 6-UCU bone-attached parallel manipulator, Chin. J. Mech. Eng, 35 (2022), 35. https://doi.org/10.1186/s10033-022-00689-1 doi: 10.1186/s10033-022-00689-1
|
| [4] |
P. Yan, H. Huang, B. Li, D. Zhou, A 5-DOF redundantly actuated parallel mechanism for large tilting five-face machining, Mech. Mach. Theory, 172 (2022), 104785. https://doi.org/10.1016/j.mechmachtheory.2022.104785 doi: 10.1016/j.mechmachtheory.2022.104785
|
| [5] |
B. Ren, Z. Zhang, Design of 4PUS-PPPS redundant parallel mechanism oriented to the visual system of flight simulator, Int. J. Intell. Robot. Appl., 5 (2021), 534–542. https://doi.org/10.1007/s41315-021-00210-2 doi: 10.1007/s41315-021-00210-2
|
| [6] |
T. Liu, Y. Hu, H. Xu, Z. Zhang, H. Li, Investigation of the vectored thruster AUVs based on 3SPS-S parallel manipulator, Appl. Ocean Res., 85 (2019), 151–161. https://doi.org/10.1016/j.apor.2019.01.025 doi: 10.1016/j.apor.2019.01.025
|
| [7] |
M. Arsenault, R. Boudreau, The synthesis of three-degree-of-freedom planar parallel mechanisms with revolute joints (3-RRR) for an optimal singularity-free workspace, J. Robot. Syst., 21 (2004), 259–274. https://doi.org/10.1002/rob.20013 doi: 10.1002/rob.20013
|
| [8] |
A. K. Dash, I. M. Chen, S. H. Yeo, G. Yang, Workspace generation and planning singularity-free path for parallel manipulators, Mech. Mach. Theory, 40 (2005), 776–805. https://doi.org/10.1016/j.mechmachtheory.2005.01.001 doi: 10.1016/j.mechmachtheory.2005.01.001
|
| [9] |
A. G. Ruiz, J. C. Santos, J. Croes, W. Desmet, M. M. da Silva, On redundancy resolution and energy consumption of kinematically redundant planar parallel manipulators, Robotica, 36 (2018), 809–821. https://doi.org/10.1017/S026357471800005X. doi: 10.1017/S026357471800005X
|
| [10] |
N. Baron, A. Philippides, N. Rojas, A novel kinematically redundant planar parallel robot manipulator with full rotatability, J. Mech. Robot., 11 (2019), 011008. https://doi.org/10.1115/1.4041698 doi: 10.1115/1.4041698
|
| [11] |
C. Cheng, W. Xu, J. Shang, Optimal distribution of the actuating torques for a redundantly actuated masticatory robot with two higher kinematic pairs, Nonlinear Dyn., 79 (2015), 1235–1255. https://doi.org/10.1007/s11071-014-1739-9 doi: 10.1007/s11071-014-1739-9
|
| [12] |
D. Liang, Y. Song, T. Sun, X. Jin, Dynamic modeling and hierarchical compound control of a novel 2-DOF flexible parallel manipulator with multiple actuation modes, Mech. Syst. Signal Process., 103 (2018), 413–439. https://doi.org/10.1016/j.ymssp.2017.10.004 doi: 10.1016/j.ymssp.2017.10.004
|
| [13] |
D. Liang, Y. Song, T. Sun, X. Jin, Rigid-flexible coupling dynamic modeling and investigation of a redundantly actuated parallel manipulator with multiple actuation modes, J. Sound Vibration, 403 (2017), 129–151. https://doi.org/10.1016/j.jsv.2017.05.022 doi: 10.1016/j.jsv.2017.05.022
|
| [14] |
Z. Chen, M. Kong, C. Ji, M. Liu, An efficient dynamic modelling approach for high-speed planar parallel manipulator with flexible links, Proc. Inst. Mech. Eng. Part C, 229 (2015), 663–678. https://doi.org/10.1177/0954406214538946 doi: 10.1177/0954406214538946
|
| [15] |
A. Rosyid, B. El-Khasawneh, Multibody dynamics of nonsymmetric planar 3PRR parallel manipulator with fully flexible links, Appl. Sci., 10 (2020), 4816. https://doi.org/10.3390/app10144816 doi: 10.3390/app10144816
|
| [16] |
L. Sheng, W. Li, Y. Wang, M. Fan, X. Yang, Dynamic model and vibration characteristics of planar 3-RRR parallel manipulator with flexible intermediate links considering exact boundary conditions, Shock Vibration, 2017 (2017), 1–13. https://doi.org/10.1155/2017/1582547 doi: 10.1155/2017/1582547
|
| [17] |
M. Burkhardt, R. Seifried, P. Eberhard, Experimental studies of control concepts for a parallel manipulator with flexible links, J. Mech. Sci. Technol., 29 (2015), 2685–2691. https://doi.org/10.1007/s12206-015-0515-1 doi: 10.1007/s12206-015-0515-1
|
| [18] |
M. Morlock, N. Meyer, M. A. Pick, R. Seifried, Real-time trajectory tracking control of a parallel robot with flexible links, Mech. Mach. Theory, 158 (2021), 104220. https://doi.org/10.1016/j.mechmachtheory.2020.104220 doi: 10.1016/j.mechmachtheory.2020.104220
|
| [19] |
F. T. Colombo, M. M. da Silva, Two hybrid model-based control strategies for a flexible parallel planar manipulator, Control Eng. Pract., 127 (2022), 105306. https://doi.org/10.1016/j.conengprac.2022.105306 doi: 10.1016/j.conengprac.2022.105306
|
| [20] | S. Karande, P. S. V. Nataraj, P. S. Gandhi, M. M. Deshpande, Control of parallel flexible five bar manipulator using QFT, in 2009 IEEE International Conference on Industrial Technology, 2009 (2009), 1–6. https://doi.org/10.1109/ICIT.2009.4939693 |
| [21] |
X. Wang, J. K. Mills, FEM dynamic model for active vibration control of flexible linkages and its application to a planar parallel manipulator, Appl. Acoust., 66 (2005), 1151–1161. https://doi.org/10.1016/j.apacoust.2005.02.009 doi: 10.1016/j.apacoust.2005.02.009
|
| [22] |
V. Sonneville, A. Cardona, O. Brüls, Geometrically exact beam finite element formulated on the special Euclidean group, Comput. Methods Appl. Mech. Eng., 268 (2014), 451–474. https://doi.org/10.1016/j.cma.2013.10.008 doi: 10.1016/j.cma.2013.10.008
|
| [23] |
G. P. Cai, C.W. Lim, Dynamics studies of a flexible hub–beam system with significant damping effect, J. Sound Vibration, 318 (2008), 1–17. https://doi.org/10.1016/j.jsv.2008.06.009 doi: 10.1016/j.jsv.2008.06.009
|
| [24] |
E. Mirzaee, M. Eghtesad, S. A. Fazelzadeh, Maneuver control and active vibration suppression of a two-link flexible arm using a hybrid variable structure/Lyapunov control design, Acta Astronaut., 67 (2010), 1218–1232. https://doi.org/10.1016/j.actaastro.2010.06.054 doi: 10.1016/j.actaastro.2010.06.054
|
| [25] |
M. Dupac, D. G. Beale, Dynamic analysis of a flexible linkage mechanism with cracks and clearance, Mech. Mach. Theory, 45 (2010), 1909–1923. https://doi.org/10.1016/j.mechmachtheory.2010.07.006 doi: 10.1016/j.mechmachtheory.2010.07.006
|
| [26] |
S. Šalinić, An improved variant of Hencky bar-chain model for buckling and bending vibration of beams with end masses and springs, Mech. Syst. Signal Process., 90 (2017), 30–43. https://doi.org/10.1016/j.ymssp.2016.12.007 doi: 10.1016/j.ymssp.2016.12.007
|
| [27] |
R. J. Theodore, A. Ghosal, Comparison of the assumed modes and finite element models for flexible multilink manipulators, Int. J. Rob. Res., 14 (1995), 91–111. https://doi.org/10.1177/027836499501400201 doi: 10.1177/027836499501400201
|
| [28] |
W. Shang, S. Cong, Nonlinear adaptive task space control for a 2-DOF redundantly actuated parallel manipulator, Nonlinear Dyn., 59 (2010), 61–72. https://doi.org/10.1007/s11071-009-9520-1 doi: 10.1007/s11071-009-9520-1
|
| [29] | A. Shabana, Vibration of Discrete and Continuous Systems, Springer International Publishing, Cham, 2019. https://doi.org/10.1007/978-3-030-04348-3 |




















Figures(21) / Tables(1)

Zhen Liu, Song Yang, Chen Cheng, Tao Ding, Ruimin Chai. Study on modeling and dynamic performance of a planar flexible parallel manipulator based on finite element method[J]. Mathematical Biosciences and Engineering, 2023, 20(1): 807-836. doi: 10.3934/mbe.2023037







 DownLoad:
DownLoad: